i början av 1960-talet började biologer undersöka hur proteiner i olika arter utvecklas på sekvensnivå (zuckerkandl 1962, margoliash 1963, Doolittle & blomb asigck 1964). De analyserade proteinerna inkluderade hemoglobin (Zuckerkandl & Pauling 1962), cytokrom C (Margoliash 1963) och fibrinopeptider (Doolittle & Blomb Acigck 1964). Dessa tidiga undersökningar ledde till en anmärkningsvärd upptäckt: det visade sig att antalet skillnader mellan proteinsekvenser av olika arter var ungefär proportionella mot tiden sedan artens divergens (Figur 1).
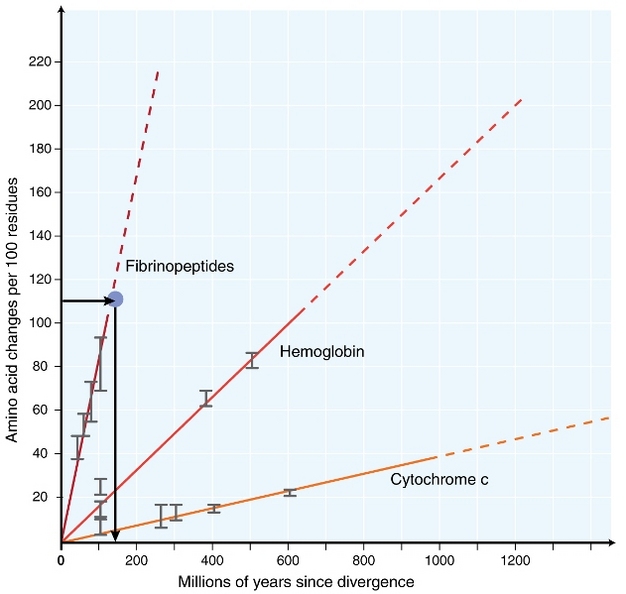
Zuckerkandl & Pauling (1965) liknade den konstanta ackumuleringen av aminosyrasubstitutioner över tiden till vanliga ”fästingar” av klockor och uppgav att ”det kan finnas en molekylär evolutionär klocka”. Således myntades termen’ molekylär klocka ’ ursprungligen för att beskriva förändringar i aminosyror som förekommer i proportionell mot tiden sedan artens divergens.
sedan dess första användning har termen ’molekylär klocka’ använts i många olika sammanhang. Numera används det ofta för att helt enkelt hänvisa till antalet förändringar, eller ’substitutioner’, ackumulerade i sekvensen av DNA eller proteiner i en given härstamning. Antalet substitutioner per en definierad tidsenhet kan beskrivas som’ hastigheten ’för molekylklockan, som i detta sammanhang motsvarar’evolutionär hastighet’. Observera att den ursprungliga konnotationen fortfarande håller i vissa fall. Till exempel, när människor säger ’vi antog en molekylär klocka’ (särskilt i fylogenetiska analyser), betyder det att antalet substitutioner antogs inträffa ständigt över tiden.
begreppet konstant molekylär klocka har extraordinära konsekvenser för evolutionära biologer. Om en konstant molekylär klocka som ursprungligen föreslogs verkligen existerade, skulle inferring timing av evolutionära händelser bli ett ganska enkelt problem (Figur 1). Det har emellertid blivit helt klart att substitutioner inte sker ständigt över tiden i olika linjer (Kumar 2005). Ändå har begreppet molekylär klocka varit extremt inflytelserikt inom molekylär evolution. En av de viktigaste ideerna inspirerade av begreppet molekylär klocka är den neutrala teorin om molekylär evolution.
molekylär klocka och neutralitet
när tanken på en konstant molekylär klocka först uppstod trodde man att den dominerande evolutionära kraften som underliggande aminosyra-eller nukleotidsubstitutioner var naturligt urval. Efter denna tankegång skulle en konstant molekylär klocka indikera att adaptiva substitutioner i olika arter uppträder ständigt över tiden. Det är dock svårt att förklara hur adaptiva substitutioner skulle inträffa på ett sådant klockliknande sätt. Teoretiskt bestäms öden för adaptiva mutationer av flera evolutionära parametrar, såsom styrkan hos den selektiva fördelen med den mutationen, storleken på den effektiva populationen och adaptiva mutationshastigheter (Kimura 1983). Dessa parametrar kommer sannolikt att skilja sig mellan arter, och även inom en art, beroende på specifika mutationer och deras interaktioner med miljöer. istället föreslog Kimura (1968, 1969) att de flesta förändringar på molekylär nivå har små funktionella konsekvenser eller är ’neutrala’. Om en mutation inte har någon konditionskonsekvens bestäms dess öde i befolkningen helt av slumpmässig slump. Detta innebär att vi inte kan förutsäga om en specifik neutral mutation så småningom kommer att fixas i befolkningen. Den hastighet med vilken neutrala substitutioner förekommer i befolkningen kan emellertid förutsägas eftersom det beror på en enda parameter, nämligen mutationshastigheten (Kimura 1968).
låt oss föreställa oss en befolkning med n antal haploida individer. Om neutrala mutationer uppträder med hastighet u per individ per generation, kommer det totala antalet mutationer i en generation att vara n gånger u. eftersom alla dessa nya mutationer är neutrala bestäms deras öde helt av en slump. Med andra ord har alla mutationer lika chans att nå fixering (vilket leder till en ’substitution’). Sannolikheten att varje ny neutral mutation kommer att nå fixering, med tanke på att en substitution inträffade, är helt enkelt 1/N. Substitutionshastigheten beräknas som antalet nya mutationer i varje generation (Nu) multiplicerat med sannolikheten varje ny mutation når fixering (1/N), vilket är lika med u. med andra ord, för neutrala mutationer är substitutionshastigheten lika med mutationshastigheten!
därför, om de flesta mutationer är neutrala (som föreslås i neutral teori) och om mutationshastigheter är konstanta över tiden, bör substitutioner också ske ständigt över tiden. Vi bör då observera klockliknande regelbundna substitutionshastigheter på molekylär nivå. Kimura (1969) ansåg således observationen av relativt konstant molekylär klocka i proteinsekvenser som starkt stöd för den neutrala teorin om molekylär evolution.
testa hur den neutrala Molekylklockan körs
enligt den neutrala teorin är frågan om substitutionshastigheter är konstanta över tiden eller inte ekvivalent med om neutrala mutationshastigheter är konstanta över tiden. Av denna anledning fokuserade många efterföljande studier på att analysera data från neutrala platser för att avgöra om neutrala mutationshastigheter verkligen är konstanta över tiden. Vi kommer kort att granska hur dessa studier genomförs innan vi diskuterar teoretiska debatter om molekylära klockor. Analyser av proteinmolekylklockor fortsatte också, men debatterna kring variation i proteinmolekylklockor skiljer sig mycket från de som rör neutrala molekylklockor och kommer inte att ingå i denna artikel. Intresserade läsare bör konsultera Gillespie (1991), Kumar (2005), Kim och Yi (2008) och Bedford et al. (2008).
de flesta empiriska analyser av neutrala molekylära klockor är beroende av satsen att neutrala mutationshastigheter kan härledas från neutrala substitutionshastigheter (Kimura 1968, 1969). I praktiken definierar varje studie en viss typ av platser i genomet som neutrala platser och jämför substitutionshastigheter för dessa platser mellan linjer.
vilka platser i genomet är verkligen neutrala kan inte bestämmas helt, men forskare kom med flera användbara proxies. Före eran med genomsekvensering var de flesta tillgängliga sekvensdata de från proteinkodande DNA-sekvenser. Studier delar ofta proteinkodande DNA-sekvenser i två typer av platser (Wu & Li 1985). Den första typen av platser inkluderar de för vilka någon förändring skulle leda till aminosyrasubstitutioner, eller ’icke-synonyma platser’. Den andra typen av platser, ’synonyma platser’, inkluderar de som kodar ’degenererade’ positioner i kodontabellen, där en förändring inte leder till en aminosyrasubstitution. TCT och TCC kodar till exempel serin. Om den tredje positionen för dessa kodon förändras kommer de fortfarande att producera samma aminosyror. Sådana substitutioner skulle vara mindre synliga för naturligt urval. Följaktligen bör molekylära klockor på synonyma platser vara närmare den neutrala molekylklockan än icke-synonyma klockor.
som sekvenseringstekniker avancerade använde vissa studier introner som proxies för neutrala platser, eftersom de inte införlivas i de mogna mRNA och därmed är mer benägna att vara neutrala (Yi et al. 2002). Sekvenser av inaktiva transponerbara element som infördes långt innan artdivergens användes också ofta (dessa kallas ofta ’förfädernas upprepningar’, t.ex. Thomas et al., 2003). Slutligen använde vissa studier icke-kodande DNA-sekvenser (alla sekvenser efter avlägsnande av proteinkodande DNA-sekvenser) extraherade från hela genominriktningar för att testa neutrala molekylära klockor (Elango et al. 2006, Huttley et al. 2007).
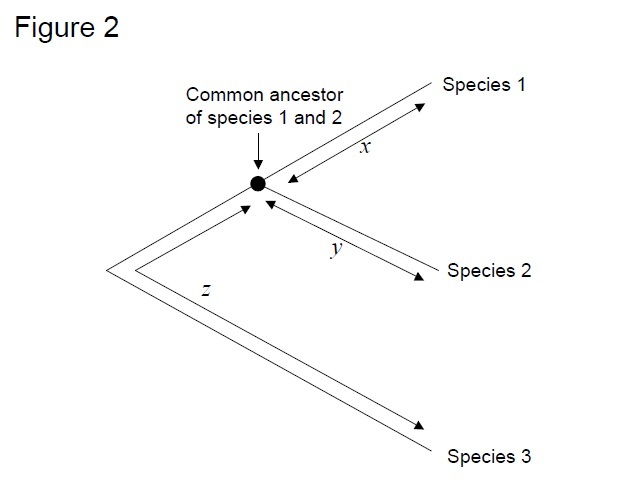
det vanligaste testet är det så kallade ”relative rate test” (Sarich & Wilson 1973). Initialt beräknades substitutionshastigheter per tidsenhet genom att dividera det totala antalet skillnader (substitutioner) mellan proteiner av olika arter med divergenstiden, beräknad från fossila register (Figur 1). Fossila register är dock inte tillgängliga för många jämförelser och är förknippade med stora felmarginaler. Det relativa hastighetstestet övervinner nödvändigheten för fossila register (Figur 2). Så länge en utgruppssekvens till de två linjerna av intresse finns, kan vi avgöra om de två grenarna följer samma eller olika molekylära klockor, utan kunskap om den absoluta tiden för divergens (Figur 2).
vilka är determinanterna för neutrala molekylära klockor?
nästan alla kontroverser i hjärtat av debatter om neutrala molekylära klockor härrör från frågan om vad de viktigaste källorna till mutationer är. Denna fråga är direkt relevant för att förstå mutationsmönster, som är den ultimata källan till evolutionär förändring och genetisk sjukdom. Att förstå hur mutationshastigheter varierar mellan linjer och inom Genom är dessutom en grundläggande fråga i jämförande genomik, som syftar till att använda sekvensjämförelser för att identifiera genomiska regioner som är funktionellt viktiga.
Så vad bestämmer neutrala mutationshastigheter? En av de viktigaste bidragsgivarna till neutrala molekylära klockor är släktspecifik variation i generationstider. Från tidigt uppfattades tanken på en konstant neutral molekylär klocka som i strid med de molekylära mekanismerna för bakteriemutation. Det har länge ansetts att de flesta mutationer härrör från fel i DNA-replikation i groddlinjer (Haldane 1947, Muller 1954). Eftersom mutationer uppstår när bakterie-DNA replikeras för nästa generation, bör de ackumuleras i proportion till antalet generationer, snarare än den absoluta tiden. Därför, om vi jämförde antalet substitutioner som har ackumulerats i två linjer sedan deras divergens, skulle linjen med längre generationstid, efter att ha genomgått färre DNA-replikationshändelser, ha färre substitutioner jämfört med linjen med den kortare generationstiden. Följaktligen bör molekylklockan springa långsammare i arter med längre generationstider. Den här tanken kallas generationstidseffekten.faktum är att generationstidseffekten först observerades i studier av primater, även före debatten om molekylär klocka. Morris Goodman, som använde immunologiska metoder för att undersöka artrelatedness vid den tiden, observerade att hastigheten med vilken vissa proteiner avviker tycktes minskas i apor, särskilt människor, jämfört med gamla världens apor (Goodman 1961, 1962, 1963). Denna effekt kallas ’hominoid hastighet avmattning’. Eftersom hominoider har längre generationstider än gamla världens apor, kan denna observation förklaras av generationstidseffekten.
Wu & Li (1985) var de första som testade generationstidseffekthypotesen med hjälp av DNA-sekvensdata. De använde data från 11 gener av primater och gnagare. Eftersom primater har en mycket längre generationstid än gnagare, bör molekylklockan vara snabbare hos gnagare jämfört med primater. De fann faktiskt att gnagare för synonyma platser visar ungefär två gånger graden av molekylär utveckling jämfört med primater (Wu & Li 1985). För icke-synonyma webbplatser hittades emellertid inte en sådan effekt. Med andra ord, den neutrala molekylklockan, men inte aminosyramolekylklockan, tickar snabbare i gnagarlinjen jämfört med primatlinjen, vilket passar bra med tanken på en generationstidseffekt.
efterföljande studier gav ytterligare stöd till avmattningen av hominoidhastigheten (Li & Tanimura 1987, Bailey et al. 1991) och hastighetsskillnaden mellan gnagaren och primatlinjerna (Gu & Li 1992, Huttley et al. 2007). Dessutom observerades en hastighetsskillnad i ännu mindre fylogenetiska skalor, särskilt i primater: till exempel går den mänskliga molekylklockan långsammare än chimpansmolekylklockan (Elango et al. 2006); priserna i New World apor är snabbare än priserna i hominoider och Old World apor (Steiper & Young 2006). De olika hastigheterna för molekylära klockor som observerats i dessa studier överensstämmer kvalitativt med generationstidseffekten.
de faktiska skillnaderna mellan linjer är emellertid inte kvantitativt förenliga med skillnaden i generationstider. Kumar & Subramanian (2002) visade att även om skillnaden i generationstider mellan primater och gnagare är mycket större än den mellan människor och gamla världens apor, är de observerade skillnaderna i molekylära klockor liknande i dessa två jämförelser. Det är värt att notera att Kumar & Subramanian (2002) använde specifika statistiska filter för att ta bort data som visade ”heterogena” substitutionsmönster, vilket kan ha orsakat en bias mot långsamt utvecklande sekvenser (Yi et al. 2002). Ändå verkar skillnaden i molekylklockorna hos primater och gnagare mycket mindre än vad som ursprungligen föreslogs av Wu & Li (1985). Till exempel Huttley et al. (2007) analyserade hela genominriktningar av flera arter inklusive opossum och visade att hastighetsskillnaden mellan eutheriska linjer och opossumlinjen (~30%) är mycket större än hastighetsskillnaden mellan mänskliga och muslinjer (~14%). Dessa exempel visar att graden av skillnader i molekylära klockor varierar avsevärt mellan olika studier på grund av skillnader i datamängder och statistiska metoder. De visar också att hastighetsskillnader mellan linjer inte helt kan redovisas av skillnaden i generationstider ensam. Klart, det finns andra bidragsgivare till neutrala molekylära klockor.
faktum är att andra livshistoriska egenskaper än generationstider verkar variera med molekylära klockor. Martin & Palumbi (1993) visade att DNA-molekylära klockor löper snabbare i arter med liten kroppsstorlek. Denna observation ledde till hypotesen att metaboliska hastigheter är viktiga determinanter för molekylära klockor. En hög metabolisk hastighet producerar ett stort antal Mutagena syreradikaler, vilket skulle öka mutationsgraden (Rand 1994). Eftersom metaboliska hastigheter och kroppsstorlek i allmänhet varierar med generationstider har det varit svårt att skilja vilka av dessa som utgör huvuddeterminanten för molekylära klockfrekvenser. Tsantes & Steiper (2009) har föreslagit, baserat på data från primater, att ålder vid första reproduktionen, snarare än kroppsstorlek, är huvuddeterminanten för molekylära klockor. Eftersom ålder vid första reproduktionen återspeglar generationstidseffekten, stöder denna studie tanken att generationstidseffekten är den viktigaste determinanten för molekylär klocka. Denna studie är dock fortfarande baserad på ett begränsat antal linjer (fyra par arter användes). Således skiljer man mellan kroppsstorlek, generationstidseffekt och metaboliska hastigheter fortfarande som en viktig fråga för att generalisera och förstå neutrala molekylära klockor. vidare har betydelsen av faktorer som inte samvarierar med generationstider, såsom DNA-metylering, uppskattats alltmer. DNA-metylering är en kemisk modifiering av genomiskt DNA som finns i olika taxa. I djurgenomer sker DNA-metylering nästan uteslutande vid cytosiner följt av guaniner (så kallade CpG). Metylerade cytosiner tenderar i sin tur att mutera snabbt till tyminer på grund av kemisk instabilitet (fågel 1980). I det mänskliga genomet förekommer mutationer orsakade av DNA-metylering mer än en storleksordning oftare än andra mutationer (Nachman & Crowell 2000, Elango et al. 2008). Eftersom mutationer orsakade av DNA-metylering förekommer i stort sett oberoende av DNA-replikation, kan sådana mutationer följa olika molekylära klockor än andra. Specifikt, istället för generationstidsberoende, kan mutationer orsakade av DNA-metylering följa en tidsberoende molekylär klocka, vilket liknar det som ursprungligen föreslogs av Zuckerkandl & Pauling (Kim et al. 2006)!
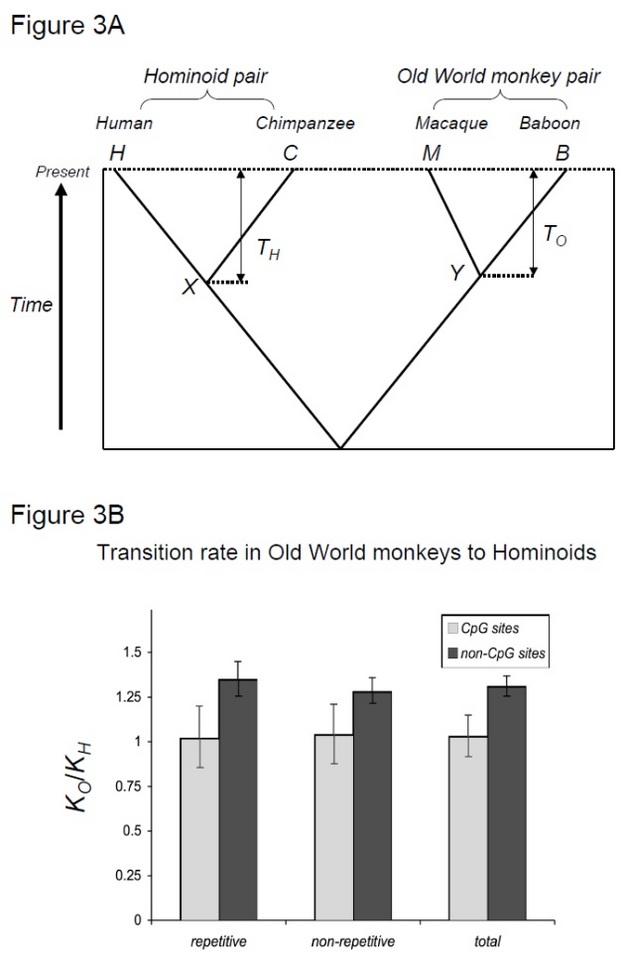
För att testa denna hypotes, Kim et al. (2006) jämförde mänsklig schimpansdivergens med makak-babiandivergens, två artpar som delar liknande divergenstider men med olika generationstider (Steiper et al. 2004). Det mänskliga schimpansparet (hominoidparet) har mycket längre generationstider jämfört med makak-babianparet (Old World monkey pair). Denna studie visade att för icke-CpG-platser ackumulerade Old World monkey-paret cirka 30% fler substitutioner, vilket kan förklaras av den ovannämnda hominoidhastighetsavmattningseffekten. Däremot visade molekylära klockor på CpG-platser liknande antal substitutioner i hominoid och Old World monkey par (Figur 3). Således existerar tidsberoende och generationstidsberoende molekylära klockor inom samma genom. Antagandet att en enda molekylär klocka kan existera för en given härstamning är inte längre giltig, eftersom de dominerande mutationskrafterna varierar mellan genomiska regioner.
slutsatser
begreppet konstant molekylär klocka föreslogs ursprungligen baserat på en begränsad mängd proteinsekvensdata. Även om efterföljande studier visade att en sådan observation inte är ett allmänt mönster i aminosyror, har den haft betydande inflytande på molekylär evolution, särskilt på utvecklingen av den neutrala teorin om molekylär evolution för DNA-sekvensdata. Efter den neutrala teorin fokuserade studier på att belysa mönster av variation i neutrala mutationshastigheter. Under de senaste decennierna har vi observerat att molekylära klockor körs i olika takt mellan linjer. Variationsgraden kan dessutom variera beroende på de olika typerna av data och specifika statistiska metoder som används. Generationstidseffekten fortsätter att hålla på en kvalitativ nivå, men är otillräcklig för att förklara kvantitativ variation av neutrala mutationshastigheter bland linjer. Livshistoriska egenskaper och icke-replikationsberoende mutationer, såsom de som orsakas av DNA-metylering, är också viktiga bidragsgivare till genomiska molekylära klockor. Faktum är att olika typer av molekylära klockor observeras även inom ett Genom, eftersom de dominerande mutationella ingångarna varierar mellan olika genomiska regioner. Således, snarare än att anta en enda neutral molekylär klocka för varje genom, bör framtida studier syfta till att avslöja variationen av genomiska neutrala molekylära klockor, för att lära sig om genomiska mutationslandskap. Sådan information är inte bara användbar för att förstå råmaterialet som styr molekylär utveckling och genetisk sjukdom, men utgör också en kritisk komponent som påverkar jämförande och funktionella genomiska analyser för att identifiera funktionella genomiska regioner.