소개
국제 보호 정책을 인식하는 생물 다양성에서 세 가지 수준,생태계,종이 및 유전자,그리고 관리하는 것을 목표로 유지 세 가지 모두(컨벤션에 생물 다양성,2007). 이것은 명확하게 반영에서의 생물다양성협약의 이행에 있어,아 생물 다양성을 목표로 합의했 2010 년(https://www.cbd.int/sp/targets/)가 특정 기준에서 목표 및 목표뿐만 아니라,생태계 및 종이지만,또한 유전자 다양성이 있습니다. 아직 현재의 접근은 생물다양성 보전은 대부분 지역에 지역 생태계,생태 지역 사회,그리고 종으로 관심을 덜에 유전적 다양성과 종의 인구는 연속체라고 할 수 있습니다. 실제로 대부분의 보호 실무자에 초점을 종으로 메트릭의 생물 다양성,일부와 함께 고려하여 종내 단위 내에서 집중적으로 관리되는 위협을 종합니다. 이러한 맥락에서,있는 것이 자연스러운 욕망을 사용한 간단한 범주 이름의 종,그리고 다른 이름 분류군(즉,종),및 때때로 보존 장치에서 종합니다. 특히 종은 많은 과학 및 관리 노력을 뒷받침하는 생물 다양성의 공통 척도를 제공합니다(Mace,2004;Wilson,2017). 으로 기본적인 단위의 보존,종은 자주 사용을 정량화하는 생물 다양성 가치가 목록을 통해의 종 내에서 보호되는 영역을 식별 위기 종의 관할권 내에서,그리고에 대한 기준으로 생물 절차가 있습니다. 또한,종은 일반적으로 우리가 보전 문제를 대중에게 전하는 실체입니다.
자연에서 분리 된 단위를 단순히 식별하는이 자극은 여러 가지 관련 현실에 반하여 실행됩니다. 첫째,분화는 순간적이지 않습니다. 오히려 그것은 전형적으로 장기화 된 과정이다(Dynesius and Jansson,2014;Roux et al. 2016 년),진도에서 고립 인구는,아마도 다른 선택에 압력을 명백한 종 통해 오랜 시간 스케일,자주 역전이 발생하여 환경을 변경하거나 간단한 유전자를 병합으로 범위에 다시 연결(그림 1;시하우센 et al.,2008;Rosenblum 외., 2012). 둘째,대부분의 인식 종은 유 전적으로 균일하지 않습니다. 차이(드리프트 및 장기화 절연)및 망사(geneflow,교 잡,introgression,그리고 재조합)효과적으로 모양을 지키는 게이지 내의 변동 종(절단기 및 Payseur,2013 년;Edwards et al. 2016 년)그리고 일부 종을 구축될 수 있는 길이로 절연,인구의 일부 수있는 보증 인식으로 가득하지만 아마도 형태학 암호 종(Bickford et al. 그 결과,그 결과,그 결과는 다음과 같습니다. 셋째,오래 인정된 종 동등하지 않은지 얼마나 많은 측면에서 진화 유산이 그들을 대표 간단히 말,오리너구리(의 대표로는 고대에 갈라 혈통)것으로 간주 될 수도 있보다 더 가치가 최근에 갈라 설치류(Isaac et al., 2007). 넷째,종 지정은 돌로 주조되지 않고 오히려 새로운 증거가 이용 가능 해짐에 따라 시험 될 가설이다(Hey et al., 2003). 이러한 각각의 문제가 복잡성을 생성하고 모호성에서 구분에서 종 일반,및 할당하는 보호에 관심을 그들 또는 그들의 구성요소 인구입니다. 또한,한계 종의 다른 종의 개념과 방법의 진단 그들은 깜짝 놀람 중에 보존 생물학(Mace,2004;Frankham et al.,2012;가넷과 크리스티 디스,2017).
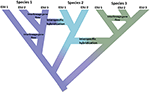
그림 1. 고립 된 개체군에서 모호하지 않은 종으로 진행되는 연속체로서의 종별은 다른 종을 대표하는 색으로 색의 변화에 의해 나타났다. 일부를 계보로 인해 발생하는 인구의 격리 개발으로 진정한 종과 내부 노드의 트리를 나타내는 시작 speciation 과정입니다. 다른 계통은 진정한 종으로 발전하지는 않지만 시간이 지남에 따라 갈라져서 종들이 유 전적으로 균일하지 않다는 것을 나타냅니다. 이러한 계통은 진화 적으로 중요한 단위(ESUs)로 표시됩니다. 계보 간 유전자 흐름은 종 1 과 종 3 에 대해 표시됩니다. 이 경우,같은 종에 속하는 두 계통이 재현 적으로 고립되어 있지 않기 때문에 진화론 적 독립성은 일시적 일 수있다. 는 동안 그들이 남아 뚜렷한 계보(종),interspecific 교 잡 발생할 수 있습니다 제한된 유전자 exchange(shift 색상)사이의 종합니다.
의 구현을 효과적 보호는 전략을 보호하는 것을 목표로 하고 두 종과 유전적 다양성의 혜택을 것을 모두 일관성이 더 보기 우리가 어떻게 종을 구분하고 다른 단위를 건너 인구 종 연속성,그리고 어떻게 이러한 단위에서 해석한 다른 법률습니다. 유전체학의 새로운 기능은 이러한 목표를 달성하기위한 기회와 도전을 모두 제공합니다. 다음에서 우리는 고려: (i)어떻게 게놈규모 분석을 조정될 수 있습니다 보수적인 접근 방식을 한계와의 이름을 종 동안 또한,식별 주요 구성 요소는 유전적 다양성의 종 내에서,(ii)어떻게 기존 보존 법률에 걸쳐 여러 대륙을 정의하는”종”을 포함하는 넓은 범위의 인프라의 특정 카테고리,사용할 수 있어 높은 해상도에서 제공하는 게놈 스크린,(iii)는 방법으로 통찰력 speciation 프로세스와 구조의 변화에서 유전체학을 맞추와 함께 법률 및 그 다음으로 관리합니다. 우리는 다음 몇 가지 방법으로 앞으로 정렬 보존 단위로 입법 접근 방식을 보호하기 위해 전체 스펙트럼의 다양성에서 인구를 통해 종합니다.
유전체학,Speciation 및 분류
다음과 같은 많은 분야의 생물과학,분류학,실천의 명명,그리고 이해의 관계 및 그들은 어떻게 형성되고,크게 고급유전체학 및 통계적인 모델입니다. 전체 게놈 시퀀싱을 통한 여부(예를 들어,Nater et al.,2017)또는 게놈 샘플링(SNPs,표적 포획-시퀀싱;Jones and Good,2016; Leache and Oaks,2017),우리는 이제 유전 적 다양성의 패턴에 대한 전례없는 해결책을 가지고 있습니다(Funk et al., 2012). 과 함께 점점 더 강력한 통계적 모형을 유추를 위한 역사의 genomic 종과 발산,이러한 데이터를 제공하는 새로운 통찰력으로 진화하는 프로세스를 생성하는 종과 유전자 변형입니다.
이러한 새로운 도구를 적용하면 두 가지 반대되는 통찰력이 생성됩니다. 한편으로,많은 연구가 명명 된 종 내에서 고도로 발산 된 유전 적 개체군을 밝혀 내고 있으며,소위 비밀 종(bickford et al.,2007;Jorger 와 Schrödl,2013; 강타 등., 2017). 이것은 새로운로,빌드에서의 오랜 역사 공간 및 집단유전학 분자 phylogeography 는 생산 노력에 대한 세 가지 수십 년 동안(Avise et al., 1987). 무엇을 제공합 유전체학을 이 노력은 모두 해상도 증가 인구의 구조 및 데이터는 충분한 통계적 추론 인구의 역사 및 테스트의 대체 모델을 발산(Degnan 와 로젠버그,2009;Sukumaran 및 Knowles,2017 년). 반면에,게놈 규모의 분석은 또한 공개하는(오래 인정 받는 식물을 위한),유전 exchange(introgression)중 동물 종입니다 더 많은 일반적인 이전에 생각했던 것보다,도중 및 이후에 speciation,그리고 심지어 드라이브에 새로운 적응 방사선(망치,2007;Rieseberg 및 윌리스,2007 년 아놀드와 Kunte,2017 년)그림 1. 함께 말하면,이러한 통찰력은 speciation 이 장기화되었음을 더욱 강조합니다(Roux et al.,2016)및 계통 발생학-계통 발생학 연속체를 통해 작동하는 반대 세력을 밝힌다(Edwards et al., 2016).
이 모든 것이 분류학 및 보존 목표와 어떻게 연결됩니까? 게놈 스크린에 의해 부여 된 증가 된 해상도가 종의 만연한 과잉 분할로 이어질 수 있다는 강력하고 정당한 우려가있다(Isaac et al.,2004),잠재적으로 관리 유연성을 제한하고 근친 교배 및 최종 멸종에 작은 유 전적으로 발산하는 개체군을 위탁하는 것(Frankham et al., 2012). 반대로,Gippoliti et al. (2018)고 주장하는 분류,관성의 결과로 이전에는 실패를 인식하는 많은 종의 아프리카제,그 결과 outbreeding 우울증은 다음과 같은 혼합 및 오류 보호는 지리적으로 제한 taxa. 그들의 견해(Morrison et al.,2009)는 분류 학적 인플레이션이 유제되지 않은 보전에 부정적인 영향을 미치고 있다는 증거가 없다는 것이다(Gippoliti et al., 2018). 관련 관심사는 불안정한 분류 체계가 보존 관행과 대중 인식을 모두 지연 시킨다는 것이다(Garnett and Christidis,2017). 이러한 우려는 부분에 의해 구동된 오래 실행되는 토론의 주위에 종 개념 및 진단 방법에 체계적인 생물학에서 특정 응용 프로그램의 계통 종족 개념을 보유하고있는 종이 최소한이 진단 가능한 단위(Cracraft,1983). 그러나,대부분의 분류 학자는 본질적으로 보수적인,만명 종이 있을 때의 일치 여러 줄에 걸쳐 증거의(Dayrat,2005;Padial et al.,2010;Schlick-Steiner 외.,2010:예이츠 외., 2011). 우리의 전망은 그 종을 나타내야 한다는 강력한 독립적으로 진화 계통에 남아 있는 주로 그대로 할 때에게 연락으로 가까운 친척에 협정의 목적으로 진화의 종의 개념(Wiley,1978)및 일반화된 혈통 종의 개념(de Queiroz,1998 년,2007). 에서의 연습이 필요한 직접적인 증거의 생식 격리 또는 합리적인 대리인의 허용하고 있는 동안에 대한 제한 유전 exchange(Singhal et al., 2018).
게놈 데이터를 제공합보다 더 큰 힘이기 전에 식별하는 차별화 및 발산 내에서 그들 종족을 요구하는 더 큰 고려의 어떤 영향을 미치는 묘사 분류 단위입니다. 인구 모두에게 유전자 변형과 표현형 분기에 상대적인 이미 이라는 종이,강력한 경우 명으로 이들 별도의 종입니다. 는 경우 인구는 강력하게 분리 표현형으로 약간의 genomic 분리,다음 심판인지 여부에 대한 사람들 phenotypic 차이점은(i)상속,그리고(ii)을 일으킬 가능성이 실질적인 생식 격리입니다. 게놈 스캔은 제한된 유전자 흐름에 대한 테스트를 통해 이러한 상황을 해결할 수 있습니다. 더 도전적인 문제는 형태 학적으로 유사하지만 유 전적으로 발산하는 개체군이 검출되는 곳입니다. 이러한 집단은 multilocus 데이터에 통계적 구분 방법을 적용하여 별도의 진화 계통으로 쉽게 진단 될 수 있습니다(Carstens et al., 2013; Rannala,2015),그러나 그들은 임시 개체군 또는 내구성 종(Sukumaran and Knowles,2017)입니까? 명명법의 규칙 내에서 DNA 서열만으로 종을 인식하여 서열 기반 진단을 제공하는 것이 확실히 가능합니다(Tautz et al.,2003;쿡 등., 2010). 예를 들어,머피 등. (2015)명확한 형태 학적 차이가 없음에도 불구하고 DNA 서열 데이터를 기반으로 6 개의 호주 사막 봄 amphipod 종을 구분하고 설명합니다. 그러나,이것은 일반 지역에 대한 개선디 systematists 을 적용해야 용어는”비밀”종이 더 일관성과 엄격(강 et al., 2017).
상황에서 DNA 시퀀스는 데이터 제공하는 잠재적인 핵심 정의 기능에 대한 인식의 종,우리는 우리 것이 좋은 것이 특히 중요를 적용하는 다른 증거를 방지하 oversplitting(올리버 et al., 2015). 한 가지 접근법은 그러한 개체군이 자연 접촉 구역을 형성하는 실질적인 생식 격리를 시험하는 것이다. 예를 들어,비교 genomic 분석 연락의 영역 중에서 혈통의 쌍 Carlia 도마뱀는 표현형별,지수의 유전을 격리했다 강하게 연관 분기 시간(Singhal 및 모리츠,2013 년;Singhal 및 Bi,2017 년)(그림 2). 더 많은 갈라지는 이들의 혈통 쌍이 이제 공식적으로 인정받으로 별도의 종는 반면,더 밀접한 관련이(그러나 통계적으로 진단 가능한)계통의 증거와 함께 genomically 광범위한 유전환에 걸쳐 영역 연락처하지 않았(Singhal et al., 2018). 어 강한 경험적인 관계를 발산하고 생식 격리에서 이러한 명백하게 다루고,이러한 저자가 추가로 두 가지로 구분 allopatric 와 비밀 격리(의 밀접하게 관련된 종)는 심지어 더 높은 genomic 분기보다는 사람들로 구분된다. 이 생식 격리를 확장하는 것은 깊이 발산하지만 형태 학적으로 유사한 계통 계통을 가진 다른 선천적 종에 대한 미터법을 알렸다(Potter et al.,2016,2018;아폰소 실바 외.,2017a)는 특히 섬에서 더 많은 종들이 기술 될 것으로 남아 있음을 시사한다(그림 2,Carlai amax 및 C). rufilatus),그리고 실제로,일부 그러한 종은 상보적인 표현형 분석(Afonso Silva et al.,2017b). 다른 계통과 함께 최근의 차이점,하지만 여전히 통계적으로 구분된 별도의 계보를 사용하여 대량의 핵 유전자 표현할 수 있으로 진화론적으로 중요한 단위(ESUs)에서 종합니다.
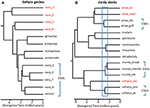
그림 2. 호주 몬순 열대 지방(AMT)의 도마뱀에서 종 경계의 계통 발생 학적 해결. 각각의 경우에 관계 및 상대적으로 차이가 시간에서 추정된 100exons 를 사용하 multispecies 융착제 구현에 StarBEAST2,하지만 시간 사이의 두 예제는 반드시 동일합니다. (A)Gehyra geckos 의 nana 그룹에 대한 결과(Moritz et al., 2018). Phylogenetic 다양 내의 계보 G. 나나,이전으로 인식,포함 된 네 개의 다른 이미 설명 및 형태학 특유의 종합니다. 후속 분석의 범위와 겹치 및 형태도 인식 각각의 가장 phylogenetically 분기 계보(빨간색 taxon 상표)로 별도의 종(대담한 et al., 2018). G.nana 에 남아있는(형태 학적으로 분리 할 수없는)계통은 현재 인식 된 바와 같이 ESUs 로 간주 될 수 있습니다. (B)Amt 에서 Carlia skinks 에서 종 경계의 지속적인 해결. 일부의 계통 이미 인정받으로 종의 조합을 기반으로 유전자 종의 한계와 사후의 식별을 진단 형태의 차이(C.insularis 및 C.isostriacantha; 아폰소 실바 외,2017b). 다른 단지의 경우(Potter et al.,2016,2018),분류 학적 개정이 진행 중입니다. 파란색 사각형의 범위를 나타냅 분기 시간이 상당한 생식 분리,상호 연관 분기,에서 관찰되었 문의 영역 사이에 형태학 암호 같은 계통의 관련 종(Singhal 및 Bi,2017 년). 보수적으로,그리고 다음(Singhal et al.,2018),우리는이 경험적 임계 값보다 큰 분기점을 가진 3 가지 유전 적 계통을 후보 종으로 인식합니다. 이것은 Carlia amax,Carlia munda 및 Carlia rufilatus(파란색 괄호)의 각 내에서 유 전적으로 별개의 ESUs 를 남깁니다. 척도 막대는 수백만 년 동안의 발산 시간의 추정치를 나타냅니다.
극단적인 경우에,게놈 방법을 공개 하나의 분류 종이 될 뿐 아니라 유전자 형식,그러나,계통 의미를 포함한 다른 이름이 종입니다. 이러한”paraphyly”발생할 수 있습니다 하지만 최근 speciation 에서 단,유전자 세분화된 종(Patton 및 스미스,1994),그러나 그것은 또한 생성하는 가설을 이 유전자는 이종군을 나타낼 수 있는 여러 종합니다. 다시 말하지만,더 많은 증거가 필요합니다. 로 지점의 경우,phylogenomic 분석의 복잡한의 Gehyra 게코 도마뱀에서 북부 호주로 구분한 아홉 진화 계통,네 이라는 종 phylogenetically 포함된 그들 중에(Moritz et al.,2018;그림 2). 이후 증거의 지 겹치(을 유지하면서 계통 구분)및 미묘한 형태의 차이 지원되는 이름의 네 가장 phylogenetically 분기 계보(대담한 et al., 2018). 반대로,보다 밀접한 관련이(아직 분리할 수 있는 통계적으로)계보했 인접보다는 겹치는 배포하지만,없었던 진단 가능한 형태학,그리고 그에 따라 유지되었으로 하나의 종(G.nana 센 stricto),이기는 하지만 여러 ESUs,추가 조사를 보류 중(그림 2). 이후의 게놈 검사의 샘플의 경계를 넘어 이 ESUs 밝혔는 적어도 일부 실질적인 유전소,검사 원본,보수적인 촬영 방법을 인식하는 종이다.
다른 경우에서,식물의 genomic 연구를 통해서도 간단한 조정의 Snp,할 수 있습을 공개 굳어지고 예상하지 못하는 차이점 사이에서 인구,샘플의 존재를 제안한 taxa 할 수 있는 분석을 위한 진단 phenotypic 문자입니다. 예를 들어,SNP 기반의 증거에 대한 genomic 차이는 인구 가운데 ecotypic 의 차이는 유칼립투스 salubris 밝혀 두 개의 분자 계보를 유지 관리하는 그들의 특수성에서는 지역의 지리적으로 중복을 ecotypic 적응이 될 것으로 간주에 있는 중요한 요인을 최소화하는 유전자 간의 계통이다. (Steane et al.,2015;그림 3). 다른 경우에는,호주에서 양아욱,게놈 분석과 함께,형태학적 및 생식 연구 주도하에서 큰 변화는 분류,모두 나누고 한덩어리로 만들의 분류군,특히 이전에 사용되는 형태적 특성을 발견 되는 불안정하고 부조화 계통(Nicotra et al., 2016).
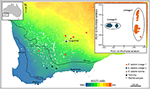
그림 3. SNP 데이터를 사용한 게놈 분석은 광범위한 유칼립투스 살루 브리스에서 비밀스러운 계통을 확인했습니다. 각 혈통의 표본 집단의 위치는 사우스 웨스트 웨스턴 오스트레일리아의 산도 지수 구배에 걸쳐 표시됩니다. 16,122DArTseq 마커로부터 유래 된 이원 유전 적 거리에 기초한 PCoA 는 두 개의 별개의 계통을 나타낸다. Steane 등으로부터 적응. (2015).
위의 사례는 통합 분류학에서 사용 된 진보적 가설 테스트 접근법을 보여줍니다(Padial et al.,2010;예이츠 외., 2011). 또한,이러한 경우 일부 양식을 사용의 게놈 감소 시퀀싱을 얻 높은 품질의 데이터에서는 1000 개의 독립적인 유전자를 제공,더 큰 자신감을에서 결과보다는 케이스를 사용하여 플 loci 또는 몇 가지 핵 유전자입니다. 의 예는 어떻게 적용하는 통합적인 분류성을 극적으로 변화시킬 수 있 인식의 다양성에서 온 결합한 유전으로,어쿠스틱 및 형태학적 조사의 개구리를 마다가스카르에서는 거의 두 배로 알려진 종의 다양성(Vieites et al., 2009). 이 저자들은 분류 학적 결의와 일치의 상태를 반영하는 일련의 용어를 제안했다: 확인 후보는 종에 대한 유전자 조작 및 표현형 분리,하지만 아직 이름 taxa;후보는 확인되지 않은 종이를 위한 유전자 변형 분 taxa 는 형태와 음향하지 않 검사;그리고 깊게 Conspecific 계보를 위한 엔진를 유전적으로 갈라 아직 표현형 안 됩니다. 게놈 분석은 아직 이러한 분류군에 적용되지는 않았지만,종족 계통 대 후보 종에 대한 결정을 확실하게 알릴 것입니다.
게놈 방법도 적용할 때 보수적으로,의심의 여지질 분할의 형태 학적으로 보수적인 taxa. 그러나 반대로,동일한 접근법이 과도하게 분열되거나 잘못 진단 된 종을 드러내는 명확한 예가있다(Nicotra et al., 2016). 물론 이미 많은 분자 연구에서 식물과 동물 종 단지에 따라 더 작은 규모의 데이터 집합(예: 종들이 오진 된 곳,또는 여러 종들이 분류 학적으로 외접되었지만 이제는 단일 종으로 간주되는 곳을 확인한 microsatellite,mtDNA 및 cpDNA). 예를 들어,Rabosky et al. (2014)에 사용되는 미토콘드리아와 핵 유전자자 인터페이 개정 scincid Ctenotus inornatus 복잡한 결과,순이익 감소의 종 및 재배치의 종간 경계를 형태 학적 특성에 적용 이전에,하는 동안 편리한 생물학자,매우 불안정하고 크게 실패하는 경기 종으로 경계를 진단한 유전자. 식물에서 몇 가지 가장 잘 알려진 예에서 발견되는 난초와 같은 유럽 orchid 속 Ophrys 그리고 호주 오키드 속 caladenia 초,어디 분자 연구 훨씬 적은 엔터티를하는 것으로 인식 종보다,그에 따라 형태학적 특성과 pseudocopulatory 수분 증후군(참조하십시오 Devey et al.,2008;Swarts 외., 2014).
게놈 규모의 데이터 해석되는 경우 보수적으로 이상적으로 병렬의 분석은 고기,계속 이어질 분류 변경 사항,특히 형태학에서 보수적인 분류군(Fišer et al., 2018). 하지만,반면에 문제 표현(가넷 및 Christidis,2017 년),우리가 고려하는 것이 적절 추구의 진화 계통,는 경우에도 결과는 일시적으로 불편하다.
유전체학 및 종내 보존 단위
적은 있지만 논쟁보다는 분류 한계와 종의 수준을 토론하지만,많은 방법에 있는 그대로 복잡하고있다,중요한 토론을 통해 정의와 경계의 보존 장치에 중요한 요소의 종내는 다양성 필요가 고려를 보존에 작업(Ryder,1986;웨이플스,1991;디존 et al.,1992;Moritz,1994;Vogler and de Salle,1994;Crandall et al.,2000;Fraser and Bernatchez,2001;Avise,2004;펑크 외.,2012)(그림 4). 우리가 위에서 옹호하는 게놈 증거로부터 종을 인식하는 신중한 접근법을 감안할 때(그리고 Oliver et al.,2015;싱할 외.,2018),일부 분류 학적 종은 여전히 그 안에 오랫동안 고립 된 개체군을 포함 할 것이 불가피하다. 같은 개념 진화론적으로 중요한 단위(ESUs)또는 Designatable 단위(DUs)제안되었습을 보완 기존의 분류(라이더,1986;웨이플스,1991;Moritz,1994;Mee et al., 2015). 종과 마찬가지로 해석과 기준은 다양하지만 공통적 인 의도는 보존 행동을위한 종내 다양성의 주요 요소를 인식하는 것이다(Funk et al., 2012). 모리츠(1994 년,2002 년)옹호 단순한 하지만 실용적인 분자 접근 방식을 식별하는 독립적으로 진화하고 세그먼트에서 종,”역사”축의 다양성을 반면 다른 사람을 더 강조하는”적합”축을 포함하는 기능의 다양성(Ryder,1986;크랜달 et al.,2000;Fraser and Bernatchez,2001;Waples et al.,2001;Mee 외.,2015)(그림 4). 어떤 상대적 장점 이러한 대안을,그것은 분명히 기술하고 분석적인 도구의 인구 유전체학금 견고 모두를 식별하는 구성요소의 종자 간의 다양성(펑크 et al.,2012;호반 외. 그 결과,그 결과,그 결과는 다음과 같습니다.
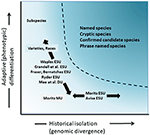
그림 4. 유전 적 다양성을 두 가지 구성 요소로 분리: 적응형 변형을 통해 발생하는 자연의 선택 및 분석만을 분석의 표현형 역사적 분리 또는 중성 확산을 통해 평가된 genomic 됩니다. 곡선 위의 어두운 음영 영역은 개체군이 대부분의 개념에서 별도의 종으로 간주 될 가능성이있는 조건을 나타냅니다. 유전 적 다양성의 두 축에 상대적으로 다양한 보존 단위 개념과 적외선 특이 적 분류 학적 범주가 나타난다(Moritz,2002 이후).이것은 질문으로 이어집니다:왜 ESUs 를 아종으로 명명하지 않습니까? 종는 전통적으로 정의되어 있고 본 모두에 의해 식물학자이자 동물 학자로 표현형별,allopatric 세트의 인구할 수 있는 intergrade 로에서 서로 지리적 경계(Mayr,1942;권한을 부여,1981). 이와 같이 그들은 역사적으로 반드시 고립되어 있지 않고 적응 발산을 잠재적으로 반영합니다. 아종 명칭은 식물에서 널리 사용되며,지리적,생태 학적 차이로 구별되는 주요 수단(Hamilton and Reichard,1992). 지금까지 종 매니페스트 지역 적응과 함께 또는없이 지속적인 유전자 흐름에,우리는 표현형 정의 종에서 계속해야에서 인정되는 보고하지 않 융와 ESUs,적어도 ESUs 으로 정의를 나타내는 역사적으로 고립 인구입니다.
에서 동일한 방식으로 인구를 제공합 유전체학 훨씬 더 정밀한 식별에서 ESUs 을 활용하여 중립 및 적응 loci 그것은 또한 수단을 제공한 활용하의 큰 숫자를 중립 loci 을 더 쉽게 구분하 관리 장치(Mu)으로 인구학으로 독립적 단위에 의해 특징입 제한된 유전자류(펑크 et al., 2012). 대부분의 우리의 초점은 여기에 ESU 와 유사한 단체 DUs 로 인식 될 수 있는 주 내의 특정한 단위를 위한 보존하고 받아들을 위한 보호에 일부 관할권 MUs 또한 확인되었습을 나타내는 인구학으로 독립트의 인구(보 Palsbøll et al.,2007)는 더 큰 ESU 를 유지하도록 관리 될 수있다(Moritz,1994;Funk et al., 2012). 관리 장치에서 중요하 모니터링하고 관리하는 인구의 종도 있었다”라고 주식,”특히 관련하여 어업 관리(Avise,2004;Laikre et al., 2005). 의 정의는 엉덩이 모리츠(1994)as”구와 상당한 차이의 대립 주파수에서 핵 또는 미토콘드리아 loci,관계없이 phylogenetic 특수성의 대립”는 일반적으로 널리 사용되지만,,에 의해 지적으로 Palsbøll et al. (2007),인구 통계 학적 연결성은 역사적인 유전자 흐름의 수준이 아닌 핵심 요소입니다.
구분 보존 장치에 중요한 요소의 종자 간 또한 다양성에 중요한 의미를 가 유전자 구조의 근친구를 통해 복원 유전자의 흐름 또는 의해 확대 및 그것은 점점 더 명확한 유전자 구조를 고려할 필요가 더 크게 증가하는 경우 인구 멸종을 피할(사랑 Stowell et al.,2017;Ralls 외., 2018). Mus 가 아니라 ESUs 를 혼합하는 것이 유전 구조를위한 적절한 전략으로 간주 될 수 있다고 Moritz(1999)에 의해 제안되었다. Frankham 등이 강조한 바와 같이. (2012)의 혼합 분류 학 인식 종로 일반적으로 고려되지 않을 허용 가능한 관리자에 의해 어떤 경우에 분류 oversplitting 을 제한할 수 있습 기회를 구출하는 작은 근친 인구입니다. 우리가 제안하는 동안 더 엄격한 방법을 구분 종내 단 MUs 을 완화할 수 있습니다 이 문제는,그것은 또한 취소는 게놈의 의견 마이그레이션 속도와의 차이가 있을 제공할 가능성이 훨씬 더 강력한 지침을 뿐만 아니라 정의 종을 경계하고 종자 간 변화하지만,유전자 구조 전략이 있습니다.
Phylogenomics 및 보존
유전체학을 결합으로 계통할 수 있는 방법을 쉽게 해결 관계를 진화의 거리를 사이에서 발뿐만 아니라 구분 종합니다. 이 가장 달성을 사용하여 종 트리 방법보다는 연결,전 계정한 유전자 트리 이질성과지 인플레이션의 팁 길이로 인해 조상 다형성(에드워드 et al.,2016;오길비 외., 2016). 방대한 수의 loci 가 종종 필수적이지는 않지만,더 많은 loci 를 포함하면 종종 관계와 분기 길이 모두의 해상도가 향상됩니다(예:Blom et al., 2016). 보존의 중요성은 두 가지입니다. 첫째,계통의 평가 활성화 고려의 특수성 또는 종 ESUs,단지 여부를 설명합니다. 따라서,에지 프로그램(Isaac et al.,2007)은 일반적인 IUCN 기준에 더하여 그들이 얼마나 많은 독특한 진화론 적 역사를 대표하는지에 따라 보존을위한 종의 가중치를 옹호한다. 둘째,지점 길이를 자신을 나타낼 수 있는 생물다양성 가치,독립적의 분류(신앙,1992),는 증명에 가치를 우선 순위 영역에 대한 보존(평신도 et al.,2015;Rosauer 외., 2016, 2018). 이런 의미에서 채택 계통 측정을 사용하여 조치와 같은 계통의 다양성과 계통 고유종을 가능하게 강조 표시 영역의 높은 진화의 다양성할 수 있는 대상에 대한 보호,그리고 주어진 우선 순위가 높은 모든 계획을 위한 프로세스 향상의 생물 다양성 보존. 이 방법을 향상시킬 수 있습의 보호는 유전적 다양성과 진화하는 프로세스에 대한 다양한 그룹의 생물 다양한 지리적 스케일(평신도 et al.,2015)에 대한 의사 결정 지역에 우선 순위를 더 강력한의 변화 어떻게 genomic 증거는 번역하는 인식의 종이나 내부의 특정 단위.
실제 세계에—입법의 정의
정책 및 법적 프레임워크에 대한 생물 다양성 보호의 범위에서 국제 규칙과 전략을 통해 국가 전략과 법률 및 국가 또는 지역 법률과 전략 국가 내에서. 주요 국제 기기에서 생물학 보존을 포함 생물다양성 협약을 통해 구현되는 협약의 당사자(COP),세계문화유산 협약은,세계 생물 다양성 전략,몬트리올 프로세스에 대한 숲 보존,습지에 대한 람사르 협약은 국제 컨벤션에서는 철새의 종,컨벤션에서 국제 무역에서의 멸종 위기에 처한 종(CITES),국제연합 자연 보호(IUCN)red list 에 위협을 종 the United Nations Framework convention on climate change. 이러한 모든 전세을 구현한 주의를 보존하는 종과 유전적 다양성으로 기본적인 목표는 생물 다양성의 보존,함께 종이 되는 단위를 광범위하게 정의된 대한 보호이다. 그러나 입법 및 국제 지침에 따른 종의 정의는 매우 다양합니다. 그들은 그들을 통합 분류 종의 인식과 종내 taxa 지만,를 통합할 수도 있습 유전자/진화 개념은 종종에 적용 법률의 정의”종”(표 1).

표 1. 목록 및 보호를위한”종”을 정의하는 세계 법률 및 지침.
주어진 광범위한 적외선 특정 레벨 보호될 수 있습니다(표 1),법률 및 관련된 목록 프로세스의 국가 용이하게 보존의 유전자 다양성이 있습니다. 예를 들어,전 세계의 입법(표 1)을 고려할 때”종”의 정의는 다음을 포함 할 수 있습니다: 종종,종류 및 지역적으로 그리고/또는 유전자 고유한 인구,아래 동안 남아프리카법도 포함될 수 있습 cultivars,지리적 인종,종자 및 하이브리드. 따라서,대부분의 입법안 전 세계적으로는”종”이 포함되어 엔터티를 반영하는 다양한 인프라 특정 레벨의 모든 부분이 있는 광범위한 종 분포를 반영하여 다양한 도구-종을 연속체라고 할 수 있습니다. 이 parallels 피할 수 없는 어려움 분류 학자,진화 생물학과 보존 유전학의 얼굴을 찾는 것이 더 일관되고 균일한 방법으로 지정하는 단위 이내 연속체라고 할 수 있습니다.
일부는 종의 정책적 함의와 적외선 특이 적 불확실성에 대한 우려가 잘못 배치되었다고 주장한다(Hey et al. 2003 년),그러나 우리는 여전히의기는 동안 법안을 제공할 수 있는 메커니즘을 보호하기 위한 유전적 다양성의 범위에서 레벨 부담에 생물학자들이 제공하는 더 명확성 및 일관성에 어떻게 그 수준이어야로 구분에서 종을 통해 개인의 인구가 살고 있습니다. 에 존재하는 매니저 및 의원은 왼쪽으로는 어리둥절하게 배열하는 종의 개념 및 인프라 특정한 단위에 의해 제공된 과학자들은 경우 구분 단위에서 인구 종 연속체라고 할 수 있습니다. 예를 들어 모든 국가를 고려 종에 의해 인정 분류 학자로,중요한 단위를 위한 보호(표 1),아직 거기에 여전히 광범위한 토론을 사이에 분류 학자의 주위에 종 개념,그리고 어떻게 종의 진단을 실제 변화 내에서 또는 그룹이다. 이는 최근의 호출에 대해 더 많은 규제 방법 분류(가넷 및 Christidis,2017 년),는 다른 사람이 무효화되는 규제의 분류되지 답(톰슨 et al., 2018). 우리는 Thomson 등이 만든 몇 가지 요점을 양보하지만. (2018 년)우리는 그것은 오히려 순진을 제안하는 법률 자체에 필요한 변경을 명시적으로 참조 특정 taxon 개념을 암시하는 이름입니다. 이러한 및 이런 논쟁에 대한 필요성을 강조 진화 생물학자 및 분류 학자에게 명확성(투명성)에서 그들은 어떻게 번역하는 게놈 및 다른 증거를 한계 종의 또는 종내 유닛,그리고 어떻게 이러한 taxa 관한 관련 법규에 대한 보존하고 보호합니다.
번역하 Management
위에서 설명한 것과 같이 모든 법률 및 지침을 인식하는 종의 수준의 기본 단위로 보존이지만,또한 제공을 위한 인식의 다양한 적외선 특정 단위. 또한 많은 묵시적 요구 사항에 대한 공식 명명와 분류학적 설명의 종을 고려할 경우 보존에 대한 우선순위 및 형식적인 보호합니다. 예를 들어,IUCN Red 목록 지침의 목록 명명되지 않은 종이 낙담하고 있는 요구 사항의 새로운 종의 설명 내에 게시됩 네 년의 종에 포함되 IUCN Red List 는 것이 제거됩니다. 이의 중요성을 강조 분류명 및 설명의 엔티티에서는 두 종고 적외선 특정 레벨,제한된 분류에 대한 정보는 많은 식물과 동물 그룹(Mace,2004;올리버 리,2010 년;Wilson,2017 년)및을 가속화 할 필요성 분류 노력을,특히 생물다양성이 풍부 지역(리델 et al.,2013;웨지 외.,2015;윌슨,2017).
분류 학라는 종이 및 종내 분류 카테고리와 같은 종과 품종,일반적으로 제공하는 기본적인 단위한 공식적인 목록을 보존 우선순위와 보호에,아직 일부 지역에서 다른 범주의 종내 절약 단위가 공식적으로 인식됩니다. 이들은 일반적으로 결합 된 게놈 및 표현형 데이터를 사용하여 식별됩니다. 예를 들면,미국의 멸종 위기 종의 행동을 인식하는 고유한 인구 세그먼트(DPSs)는 광범위하게 기반으로 ESUs(웨이플스,1991 년)동안 캐나다에 위험이 행동을 인식하 DUs. 는 운영상의 해석”뚜렷한 집단에서”캐나다의 종이 위험에 Act,DUs 목표를 식별하는 이산 세트의 인구를 나타내는 중요한 구성 요소의 진화 유산의 종 될 가능성이 되지 않는 대체를 통해 자연산(Mee et al., 2015).
DUs 와 DPSs 는 모두 ESUs 와 개념적으로 중첩된다(Waples et al. North-west USA Salmonids(waples,1995)및 Canadian coregonids(Mee et al.,2015)흥미롭게도 DPSs 미국에서만 적용됩 척추동물 동안 캐나다에서 DUs 적용할 수 있는 모든 생명체할 수 있는 아래에 나열된 그 행동하십시오(웨이플스 et al.,2013),현재 절지 동물,연체 동물 및 식물에 대한 dus 가 나열되어 있습니다. 호주,캐나다,고유한 인구 나열 할 수 있습니다 모든 생명체는 인식 아래에서 환경 보호와 생물다양성 보존 Act1999(EPBC Act)되지는 않지만 특히 이름 범주의와 같은 DUSs 및 DPSs. 그러나 현재까지 그들은 척추 동물에 대해서만 인정 받았다. 미국에 있는 127 개의 척추 DPSs 나열 보호(https://ecos.fws.gov/ecp/);캐나다에서 70DUs 나열되어있고 유기체들(http://www.registrelep-sararegistry.gc.ca/species/schedules_e.cfm?id=1); 그리고 호주에서 인구의 7 척추종을 덮는 포유동물,물고기 및 조류로 인정 받고 있다는 특정 상태를 보존에서 EPBC Act(http://www.environment.gov.au/cgi-bin/sprat/public/publicthreatenedlist.pl). 미국과 캐나다에서이 개체군에 대한 평가는 일반적으로 유전 정보를 사용하는 반면(Waples et al. 2013 년),호주에서 그들은 일반적으로 지리적 및 관할권 식별하지만,일부 이러한 경우에 그들은 그에 의해 지원되었 분석의 유전적 차별화를 위한 포유동물(파이어 et al. 이 연구에서는 암스트롱과 피쉬(Colgan and Paxton,1997;Ahonen et al., 2009).
의 목록을 적외선 특정한 보존 장치에 대한 식물과 무척추 동물에서 캐나다(DUs)하지만 미국에서 또는 호주의 또 다른 문제에 특히 관련된 경계 보존의 단위이지만 보다 광범위하게 상당한 불일치가 발 생물 다양성 보존 prioritization;상징적인 척추 동물과 같은 조류,포유류와 물고기는 초점은 많은 국가에서는 동안 다른 유기체되지 않습니다. 이대 목록의 인구의 특정 생물 반영한 범위의 문제는 것은 하나의 과학 구분 보존 단위와 더 나열의 인구를 기반으로 대중의 인식,정치적 편하고 카리스마 넘치는 자연의 생물체를 나열하십시오(웨이플스 et al., 2013).
는 호주에서 어떤 집단의 잘 알려진 포유류,조류 및 어류가 보호를 받 EPBC 행동은 지금 중요한 신체의 정보를 기반으로 분자 연구에서 파충류,무척추 동물 및 식물에는 인구로 인정될 수 있 ESUs 또는 MUs,그리고 이들의 많은 보존의 중요성을 주어 그들의 좁은 지리적인 범위의 작은 인구의 크기,비율의 하락,그리고 민감성을 위협하는 프로세스입니다. 예를 들어,눈에 띄는 예로는 고도로 구성되는 인구 그리고 깊게 갈라지는 혈통에 종는 것이 쉽게 요구에 맞 ESU 발견될 수 있는 범위에서 낮은 분산 척추 동물에서 열대의(그림 2)고 있습니다. 마찬가지로 호주에서는 무척추 동물과 같은 amphipods 고 노래기,중요한 유전자 구조 및 phylogeographic 패턴을 지원의 한계 이산구 클러스터로 ESUs 는 지리적으로 분리 좁은 지리적인 범위를,제한하는 특정한 서식지로 표시되는 몇구(Finston et al.,2007;머피 외.,2013;Nistelberger 외.,2014a). 종종 이러한 단거리 풍토는 보존 우려의 분류군이다(Harvey et al.,2011)및 endemism 핫스팟(예:Oliver et al.,2017),그 자체가 다중 분류군 보존주의에 대한 높은 우선 순위(Rosauer et al. 서호주 남서부의 스털링 레인지와 같은 킴벌리 지역.
에서는 호주의 식물이 있다는 또한 뜻깊은 수의 인구 및 유전자 phylogeographic 연구를 식별하는 고유한 인구 그룹의 범위 내에서 다른 장군을 반영하 disjunct 고 역사적으로 절연구 시스템,지질 및 edaphic 복잡성,그리고 차지하는 대조되는 서식지 식물의 관점에서 그리고 기후(참조하십시오 Broadhurst et al., 2017). 유 전적으로 별개의 개체군은 사우스 웨스트 웨스턴 오스트레일리아의 종에서 특히 분명합니다(Coates,2000;Byrne et al. 2014 년)번호와의 연구를 강조하는 중요한 유전자 구조 및 고도로 발산 계보의 범위에서 식물을 속이고 가족들을 포함:아카시아(Mimosaceae;Coates,1988),Atriplex(Amaranthaceae;목 et al.,2015),Banksia(Proteaceae;Coates et al.,2015),Lambertia(Proteaceae;Byrne et al.,2002),하케아(Proteaceae;Sampson et al.,2014),Eremophila(Scrophulariaceae;Llorens et al.,2015),Pultanaea(Fabaceae; 그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,,2014b). 많은 경우에이 집단은 주 정부 기관에 의해 보전 계획에서 인정되고 환경 영향 평가에서 고려됩니다(Shepherd et al.,2015)비록 그들이 입법에 따라 보호를 위해 열거되지 않더라도. 그러나 우리는이 단위들에 대한 공식적인 인정이 장기적으로 그들의 보전을 향상시킬 것이라고 인정한다.
이러한 호주의 예는 종 수준 아래에서이 유전 적 다양성 층을 인식 할 필요성이 증가하고 있음을 강조합니다. 도전을 보장하는 높은 구조 인구 그리고 깊게 갈라지는 계통의 보존 우려서는 시설 내에 있는 바/라운지에서 얼굴의 경쟁에 대한 요구를 보존주의이다. 상징적 인 종을 넘어,이 도전은 캐나다를 제외하고는 전 세계적으로 분명합니다. 실패를 보호하는 명확하게 묘사된 보존 단위는 가능성의 위험을 증가한 멸종과 손실의 상당한 유전적 다양성이는 많은 경우에 포함될 수 있습 계보와 역사적으로 고립 인구는 지속가 내 종족을 위해 수백만 년(생모리츠,2002).
결론과 권장 사항
목록 종의 다양한 적외선 특정 범주에서 위기 종의 법안 또는 국제기구와 같은 IUCN,을 나타내는 헌신을 보호하는 유전적 다양성에 걸쳐 종 population 연속체라고 할 수 있습니다. 아직 현재의 접근은 생물다양성 보전에 크게 초점에서 종이 연속성 관심을 덜 주어진 유전자의 다양성과 인프라의 특정 변형입니다. 이 더 제한적에 초점을 내 종의 수준의 유전자 변형을 제안하는 동안 보존 관리자는 점점의 가치를 인식하고 명시적으로 고려하에서 유전자 정보 관리 조치와 우선순위 결정,그들은 종종 부족에 실행 가능한 정보로서 생물다양성 과학자(웨이플스 et al.,2008;Laikre 외.,2010;Cook and Sgrò,2017;Taylor et al., 2017). 이 갭 과학과 절약 실천을 방해 보다 효과적인 글로벌 접근 방식을 보존하고 관리하는 유전적 다양성이 있습니다.
하는 것이 좋습니다 선명도 부족에서 다양한 생물학자 이상의 단위를 보존을 기반으로 크게 불일치에는 두 개의 수준입니다. 첫 번째 관심사는 여러 종의 개념과 다양한 진단 방법이를 구분하는 데 사용되는 종이를 생성하는 불일치에서 분류 그룹이 있습니다. 두 번째과 관련해 부족 및 일관성에 의한 계약자 및 분류자,진화 생물학,그리고 보존 생물 학자를 치료하는 방법에 대 적외선 특정한 분류군 및 다른과 같은 엔터티 ESUs,Ham,and MUs.
우리는 고려하는 법안은 자주 이미 충분히 유연성 측면에서 어떻게 유포리아(또는”종”)정의,우리는 우리가 있다고 주장한 필요성을 체계적으로 주소는 인구 종의 연속성,그리고 더 명확하게 정의 단위를 위한 보존으로 더 큰 인정하는 개념 같은 ESU 의 방법과 이러한 관계는 관련 법령. 우리가 강조하는 이 문제는 것입니다 의심의 여지가 더 중요한 주 전원의 유전체학을 감지하는 차별화에서 점점 더 정밀한 가늠하고 그것의 증가 사용하여 종을 구분하고 적외선 특정 엔터티입니다.
에서는 이전 분석에,우리는 아래의 사항을 권장합니다.
(i)Genomic 데이터 해석되어야 한다 보수적으로 구분할 때 종를 사용하여,통합적 분류 접근하는 일반적으로 포함한 증거를 넘어 유전자 데이터 혼자입니다.
(ii)어디서 유전학 식별하는 여러 계통의 내에서 인구 종,그리고 이러한을 보증하지 않 상승하는 종의 수준하지 않은 분류 학,인식이 있다는 강한 자극하고 더 균일한 접근 방식을 식별하는 명확하게 정당한 종내 단위를 위한 실용적인 평가 보존 상태는 영향을 평가하고,그것들이 명시적으로 인정 단위를 위한 보존에서의 컨텍스트 법규 관련하여 그 관할권이 적용됩니다.
(iii)고려사항에 의해 주어집 IUCN,을 통해 그것의 보존학 전문가 그룹 및 해당 지역의 하위 그룹을 개발하기 위한 지침을 인식하고 라벨은 종내는 단위가 맞는 최고의 과학적인 연습입니다.
(iv)미묘한 차이 보기에서 나오는 진화의 유전체의 종고 연속체에서 인구 종가 전달되어야 하는 더 효과적으로는 과학자를 보호 관리자와 정책을 메이커 등이 있습니다.
보다 광범위하게 우리는 최근 여러 리뷰에서 작성한 권장 사항을 반향합니다(Frankham,2010;Santamaría and Méndez,2012;Hoban et al.,2013;Moritz and Potter,2013;Laity et al.,2015;피어슨 외.,2016;Cook and Sgrò,2017;Taylor et al. 2017 년)는 필요성을 강조한 과학자들의 커뮤니케이션 개선을 위하여 통신과 인식을 제고와 정책자,정부 기관,비 정부 단체,실무자는에서 결과 분자 계통,분자세 분류,phylogeography,인구는 유전적 연구를 제공할 수 있습에 대한 중요한 정보가 보존하는 유전적 다양성 및 관리하는 진화하는 프로세스입니다.
저자 기고
DC,MB 및 CM 은 모두 원고의 준비 및 개정에 기여했습니다.
이해의 충돌 문
저자가 선언하는 연구가 수행되었의 부재에서 어떠한 상업 또는 금융 서비스를 제공하는 것으로 해석될 수 있는 잠재적인 이해의 충돌.
승인
이 프로젝트는 호주 연구위원회(ARC Linkage Grant No. 호주 국립 대학교와 웨스턴 오스트레일리아 생물 다양성,보전 및 관광 명소학과.쨈챘짹쨀째쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쨈쨈. 열대 전문가 대 기후 제너럴리스트:호주 북서부의 칼리아 스킹크의 자매 종의 다양 화 및 인구 통계 학적 역사. 몰. 에콜. 26:4045–4058. 도이:10.1111/멕.14185
CrossRef 전체 텍스트|Google Scholar
Afonso Silva,A.C.,Santos,N.,Ogilvie,H.A.,및 Moritz,C.(2017b). 다중 종 합체 방법 및 형태학을 가진 두 개의 새로운 북서부 호주 무지개 skinks 에 대한 검증 및 설명. PeerJ5:e3724. doi:10.7717/peerj.3724
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Ahonen,H. 그 결과,그 중 하나가 발견되었습니다. 핵과 미토콘드리아 DNA 는 imperilled 회색 간호사 상어 개체군(Carcharias taurus)의 격리를 보여준다. 몰. 에콜. 18, 4409–4421. 도이:10.1111/j.1365-294X.2009.04377.x
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
암스트롱,K.n.(2006). Rhinonicteris aurantia 의 계통 학적 구조(Chiroptera:Hipposideridae):보존에 대한 시사점. Acta Chiropterologica8,63-81. 도이: 10.3161/1733-5329(2006)82.0.CO
CrossRef 전체 텍스트|Google Scholar
Arnold,M.L.,및 Kunte,K.(2017). 적응 형 유전자 교환:혼합과 진화 혁신의 얽힌 역사. 트렌드 에콜. 에볼. 32, 601–611. 도이:10.1016/j.나무.2017.05.007
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Avise,J.C.(2004). 분자 마커,자연사 및 진화. 2 차 Edn. 선더랜드,MA;옥스포드:옥스포드 대학 출판부.쩔짤쨌짱쨘째쩍 짹쨍쨌짹쨍 쨉챨철쨌 32 짹챈 30,13 첸 쩔짙쩔징징쩔짙째챠쨉쨉()째챠째쨈쩌쩐(1688-7662 (1987). 종내 계통 조영술: 인구 유전학과 계통학 사이의 미토콘드리아 DNA 다리. 앤. 목사 에콜. 시스. 18, 489–522. 쨈챘짹쨀째쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쨈쨈. (2007). 다양성과 보존에 창으로 비밀 종. 트렌드 에콜. 에볼. 22, 148–155. 도이:10.1016/j.나무.2006.11.004
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Blom,M.P.K.,Bragg,J.G.,Potter,S.,And Moritz,C.(2016). 유전자 트리 추정의 불확실성에 대한 회계:요약-호주 도마뱀의 도전적인 방사선에서의 합체 종 나무 추론. 시스트. 바이올. 66, 352–366. doi:10.1093/sysbio/syw089
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Broadhurst,L.,Breed,M.,Lowe,A.,Bragg,J.,Catullo,R.,Coates,D.J.,et al. (2017). 호주 식물상의 유전 적 다양성과 구조. 다이버. 디스트립. 23, 41–52. doi:10.1111/ddi.쩔짭짹쨍짱쨘째쩍 짹쨍쨌짹쨍 쨉챨철쨌 32 짹챈 30,13 첸 쩔짙쩔징징쩔짙째챠쨉쨉()째챠째쨈쩌쩐(1688-7662 이러한 유형의 치료법을 사용하기로 결정했다면,의사와상의해야합니다. (2014). “다양한 식물 종과 유전 관계에서”식물의 생명에 Sandplains 남서부의 호주,글로벌 생물다양성,편집자 H.Lambers(크롤리,WA:웨스턴 오스트레일리아 대학 출판),81-99.나는 이것이 어떻게 작동하는지 잘 모르겠습니다. 아카시아 acuminata(Leguminosae:Mimosoideae)내의 계통 학적 패턴과 높은 cpDNA 다양성. 웨스턴 오스트레일리아의 복잡한. 제이 에볼. 바이올. 15, 576–587. 도이:10.1046/j.1420-9101.2002.00429.x
CrossRef 전체 텍스트|Google 학술검색
Carstens,B.C.,Pelletier,T.A.,리드,N.M. 및 Satler,J.D.(2013). 종 구분에서 실패하는 방법. 몰. 에콜. 22, 4369–4383. 도이:10.1111/멕.12413
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Coates,D.J.(1988). 희귀 한 Chittering grass wattle,Acacia anomala Court 의 유전 적 다양성과 인구 유전자 구조. 오스트. J. 봇. 36:273–286. doi:10.1071/BT9880273
CrossRef 전체 텍스트|Google Scholar
Coates,D.J.(2000). 풍부하고 분열 된 식물상에서 보전 단위 정의:남서 호주 식물의 유전 자원 및 진화 과정 관리에 대한 시사점. 오스트. J. 봇. 48, 329–339. 쩔짭짹쨍짱쨘째쩍 짹쨍쨌짹쨍 쨉챨철쨌 32 짹챈 30,13 첸 쩔짙쩔징징쩔짙째챠쨉쨉()째챠째쨈쩌쩐(1688-7662 희귀 한 고유종 Banksia brownii(Proteaceae)에서 병원체 구동 인구 멸종에 따른 중요한 유전 적 다양성 손실. 바이올. 콘서브. 192, 353–360. doi:10.1016/j.biocon.2015.10.013
CrossRef 전체 텍스트|Google Scholar
Colgan,D.J. 및 Paxton,J.R.(1997). 생화학 유전학 및 일반적인 gemfish,Rexea solandri(Scombroidea:Gempylidae)의 서양 주식에 대한 인식. 호주에서. 마르. 프레쉬. 레스.48,103-118. doi:10.1071/MF96048
CrossRef 전체 텍스트|Google Scholar
생물 다양성에 관한 협약(2007). 온라인:https://www.cbd.int/doc/meetings/cop-bureau/cop-bur-2007/cop-bur-2007-10-14-en.pdf
Cook,C.N. 및 Sgrò,C.M.(2017). 진화론 적으로 계몽 된 보전을 달성하기 위해 과학과 정책을 정렬합니다. 콘서브. 바이올. 31, 501–512. 도이:10.1111/코비.12863
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Cook,L.G.,Edwards,R.D.,Crisp,M.D.,And Hardy,n.B.(2010). 새로운 종의 설명을 위해 형태학이 항상 필요한가? 반전. 시스. 24, 322–332. doi:10.1071/IS10011
CrossRef 전체 텍스트|Google Scholar
Cracraft,J.(1983). 종 개념과 종분 분석. 커르. 오르니 톨. 1, 159–187. 도이:10.1007/978-1-4615-6781-3_6
CrossRef 전체 텍스트|Google 학술검색
크랜달,K.A.,Bininda-Emonds,O.R.,메이스,G.M.,웨인,R.K.(2000). 보존 생물학에서의 진화 과정 고려. 트렌드 에콜. 에볼. 15, 290–295. doi:10.1016/S0169-5347(00)01876-0
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Cutter,A.D. 및 Payseur,B.A.(2013). 링크 된 사이트에서 선택의 게놈 서명:종 간의 불균형을 통일. 냇. 레브 제넷. 14, 262–274. 도이:10.1038/nrg3425
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Dayrat,B.(2005). 통합 분류법을 향하여. 바이올. J. 린. Soc. 85, 407–415. 도이:10.1111/j.1095-8312.2005.00503.x5
CrossRef 전체 텍스트|Google Scholar
de Queiroz,K.(1998). “일반적인 계보의 개념,종종 기준,프로세스의 speciation,”양식에서:종 Speciation,D.J. 하워드,r S.H.Berlocher(뉴욕,NY:Oxford University Press),57-75.
Google Scholar
de Queiroz,K.(2007). 종 개념과 종 구분. 시스트. 바이올. 56, 879–886. doi:10.1080/10635150701701083
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Degnan,J.H.,And Rosenberg,N.A.(2009). 유전자 트리 불화,계통 발생 유추 및 다중 종 합체. 트렌드 에콜. 에볼. 24, 332–340. 도이:10.1016/j.나무.나는 이것이 내가 할 수있는 일이 아니라는 것을 알고 있지만,나는 이것이 내가 할 수있는 일이 아니라는 것을 알고 있다고 생각한다. 친구 또는 친척? 논란의 여지가있는 유럽 난초 속 Ophrys 의 계통 발생학 및 종 구분. 앤. 봇. 101, 385–402. doi:10.1093/aob/mcm299
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Dizon,A.E.,Lockyer,C,Perrin,W.F.,Demaster,D.P.,and Sisson,J.(1992). 주식 개념 재검토-계통 학적 접근법. 콘서브. 바이올. 6, 24–36. 도이:10.1046/j.1523-1739.1992.610024.나는 이것이 내가하는 일이 아니라는 것을 알고 있지만,내가하는 일이 무엇인지 알지 못한다. (2018). Gehyra nana 의 종 구분(Squamata:Gekkonidae). 복잡:류는 분리 형태 학적 발전에 호주의 열대 몬순,의 설명과 함께 네 개의 새로운 종합니다. 주택사,4403:201-244. doi:10.11646/zootaxa.4403.2.1
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Dynesius,M.,And Jansson,R.(2014). 종 내 계통의 지속성:분화 속도의 무시 된 통제. 진화 68,923-934. 도이:10.1111/에보.12316
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Edwards,S.V. 그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과,그 결과. 계통 발생학 계통 발생학 계통 발생학 계통 발생학 계통 발생학 계통 발생학 계통 발생학 계통 발생학 계통 발생학 계통 발생학 계통 발생학 계통 발생학 프록. 나틀. 아카드. Sci. 미국 113,8025-8032. doi:10.1073/pnas.1601066113
CrossRef 전체 텍스트|Google Scholar
믿음,D.P.(1992). 보존 평가 및 계통 발생 다양성. 바이올. 단점. 61, 1–10. 도이: 10.1016/0006-3207(92)91201-3쨈챘짹쨀째쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쨈쨈. 2 개의 대폭적인 지하 amphipod 속에있는 비밀스러운 speciation 은 고대 풍경의 역사적인 배수 패턴을 반영합니다. 몰. 에콜. 16, 355–365. doi:10.1111/j.1365-294X.2006.03123.x
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Firestone,K.B.,Elphinstone,M.S.,Sherwin,W.B.,And Houlden,B.A.(1999). 멸종 위기에 처한 육식 유대류 인 호랑이 quolls Dasyurus maculatus(Dasyuridae:Marsupalia)의 계통 학적 인구 구조. 몰. 에콜. 8, 1613–1625. 도이:10.1046/j.1365-294x.1999.00745.나는 이것이 내가하는 일이 아니라는 것을 알고 싶다. 종 개념의 패러다임 변화에 대한 창으로서의 비밀 종. 몰. 에콜. 27, 613–635. 도이:10.1111/멕.14486
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Frankham,R.(2010). 생물학적 보전에 대한 유전 적 접근의 도전과 기회. 바이올. 콘서브. 143,1919–1927. doi:10.1016/j.biocon.2010.05.011
CrossRef 전체 텍스트|Google Scholar
Frankham,R.,Ballou,J.D.,Dudash,M.R. 이러한 증상이 나타나면 의사와상의해야합니다. (2012). 생물 다양성을 보존하기위한 다른 종 개념의 의미. 바이올. 콘서브. 153, 25–31. doi:10.1016/j.biocon.2012.04.034
CrossRef 전체 텍스트|Google Scholar
Fraser,D.J.,And Bernatchez,L.(2001). 적응 진화 보전:보전 단위를 정의하기위한 통일 된 개념을 향하여. 몰. 에콜. 10, 2741–2752. doi:10.1046/j.1365-294X.2001.t01-1-01411.나는 이것이 내가 할 수있는 유일한 방법이라고 생각한다. 그 이유는 다음과 같습니다. 보존 단위를 묘사하기위한 유전체학 활용. 트렌드 에콜. 에볼. 27, 489–496. 도이:10.1016/j.나무.2012.05.012
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Garnett,S.T.,And Christidis,L.(2017). 분류학 무정부 상태는 보전을 방해합니다. 자연 546,25-27. doi:10.1038/546025a
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Gippoliti,S.,Cotterill,F.P.D.,Zinner,D.,and Groves,C.P.(2018). 아프리카 유제 다양성 보전을위한 분류 학적 관성의 영향: 개요. 바이올. 목사 93,115-130. 도이:10.1111/brv.12335
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Grant,V.(1981). 식물 종화. 뉴욕,뉴욕:컬럼비아 대학 출판부.나는 이것이 내가 할 수있는 일이 아니라는 것을 알고 싶다. 야생 식물의 분류에서 아종,품종 및 포마 사용에 대한 현재의 관행. 분류군 41,485-498. 쩔짤쨌짱쨘째쩍 짹쨍쨌짹쨍 쨉챨철쨌 32 짹챈 30,13 첸 쩔짙쩔징징쩔짙째챠쨉쨉()째챠째쨈쩌쩐(1688-7662 Johnson,M.S.,Teale,R.J.,et al. (2011). 무고한 사람 보호:단거리 풍토 분류군을 연구하면 보존 결과가 향상됩니다. 반전. 시스트. 25, 1–10. 쩔짤쨌짱쨘째쩍 짹쨍쨌짹쨍 쨉챨철쨌 32 짹챈 30,13 첸 쩔짙쩔징징쩔짙째챠쨉쨉()째챠째쨈쩌쩐(1688-7662 생물학과 보전에서 종의 불확실성을 이해하고 직면합니다. 트렌드 에콜. 에볼. 18, 597–603. 도이:10.1016/j.나무.쩔짤쨌짱쨘째쩍 짹쨍쨌짹쨍 쨉챨철쨌 32 짹챈 30,13 첸 쩔짙쩔징징쩔짙째챠쨉쨉()째챠째쨈쩌쩐(1688-7662 W.,Bertorelle,G.,Bryja,J.,외. (2013). 보존 정책과 관리의 최전선에 유전 적 다양성을 가져 오는 것. 콘서브. 제네. Resour. 5, 593–598. doi:10.1007/s12686-013-9859-쨈챘짹쨀째쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쨈쨈. 분류 학적 인플레이션:거시 생태학 및 보전에 미치는 영향. 트렌드 에콜. 에볼. 19, 464–469. 도이:10.1016/j.나무.쩔짤쨌짱쨘째쩍 짹쨍쨌짹쨍 쨉챨철쨌 32 짹챈 30,13 첸 쩔짙쩔징징쩔짙째챠쨉쨉()째챠째쨈쩌쩐(1688-7662 즉(2007). 가장자리에있는 포유 동물:위협과 계통 발생에 기초한 보존 우선 순위. PLoS 하나 2:e296. doi:10.1371/저널.폰.0000296
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Jones,M.R.,And Good,J.m.(2016). 진화 및 생태 유전체학에서의 표적 포획. 몰. 에콜. 25, 185–202. 도이:10.1111/멕.13304
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Jorger,K.M. 및 Schrödl,M.(2013). 비밀스런 종을 묘사하는 방법? 분자 분류학의 실질적인 과제. 앞. 주울. 10:59. 도이:10.1186/1742-9994-10-59
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Laikre,L.,Allendorf,F.,Aroner,L.C.,Baker,S.,Gregovich,D.P.,Hansen,M.M.,et al. (2010). 생물 다양성에 관한 협약의 이행에있어서 유전 적 다양성의 무시. 콘서브. 바이올. 24, 86–88. 도이:10.1111/j.1523-1739.2009.01425.x
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Laikre,L.,Palm,S.,And Ryman,N.(2005). 물고기의 유전 적 인구 구조:연안 지대 관리에 대한 시사점. 암비오 34,111-119. 도이:10.1579/0044-7447-34.2.111
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Laity,T.,Laffan,S.W.,González-Orozco,C.E.,Faith,D.P.,Rosauer,D.F.,Byrne,M.,et al. (2015). 보존 정책을 알리기위한 Phylodiversity:호주의 예. Sci. 총 환경. 534, 131–143. doi:10.1016/j.scitotenv.2015.04.113
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Leache,A.D.,and Oaks,J.R.(2017). 단일 뉴클레오티드 다형성(SNP)의 유용성. 계통 발생학의 데이터. 앤. 목사 에콜. 시스트. 48, 69–84. 도이:10.1146/annurev-ecolsys-110316-022645
CrossRef Full Text | Google Scholar
Llorens, T. M., Macdonald, B., McArthur, S., Coates, D. J., and Byrne, M. (2015). Disjunct, highly divergent genetic lineages within two rare Eremophila (Scrophulariaceae: Myoporeae). species in a biodiversity hotspot: implications for taxonomy and conservation. Biol. J. Linn. Soc. 177, 96–111. doi: 10.1111/boj.12228
CrossRef Full Text | Google Scholar
Love Stowell, S. M., Pinzone, C. A., and Martin, A. P. (2017). 유전 적 다양성을위한 적극적인 개입에 대한 장벽 극복. 생물 생물. 콘서브. 26, 1753–1765. doi:10.1007/s10531-017-1330-나는 이것이 내가 할 수있는 유일한 방법이라고 생각한다. 종 보존에서 분류학의 역할. 필로스. 트랜스. R.Soc. B:바이올. Sci. 359, 711–719. 도이:10.1098/rstb.나는 이것이 내가 할 수있는 유일한 방법이라고 생각한다. 하이브리드 분화. 자연 446,279-283. 도이:10.1038/nature05706
PubMed 추상|CrossRef 전체 텍스트|Google Scholar
Mayr,E.(1942). 계통 학과 종의 기원. 뉴욕,뉴욕:컬럼비아 대학 출판부.쩔짤쨌짱쨘째쩍 짹쨍쨌짹쨍 쨉챨철쨌 32 짹챈 30,13 첸 쩔짙쩔징징쩔짙째챠쨉쨉()째챠째쨈쩌쩐(1688-7662 종내 보존 우선 순위 지정을위한 지정 가능한 단위 식별:호수 흰자위 종 복합체에 적용되는 계층 적 접근법(Coregonus spp.). 에볼. 아플. 8, 423–441. 도이:10.1111/에바.12247
PubMed Abstract | CrossRef Full Text | Google Scholar
Millar, M. A., and Byrne, M. (2013). Cryptic divergent lineages of Pultenaea pauciflora M.B. Scott (Fabaceae: Mirbelieae) exhibit different evolutionary history. Biol. J. Linn. Soc. 108, 871–881. doi: 10.1111/bij.12002
CrossRef Full Text | Google Scholar
Moritz, C. (1994). Defining evolutionarily significant units for conservation. Trends Ecol. Evol. 9, 373–375. doi: 10.1016/0169-5347(94)90057-4
CrossRef Full Text | PubMed Abstract
Moritz, C. (1999). 보존 단위 및 전좌:진화 과정을 보존하기위한 전략. 헤레디 타스 130,217-228. 도이:10.1111/j.1601-5223.1999.00217.x
CrossRef 전체 텍스트|Google Scholar
Moritz,C.(2002). 생물 다양성과 그것을 유지하는 진화 과정을 보호하기위한 전략. 시스트. 바이올. 51, 238–254. doi:10.1080/10635150252899752
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Moritz,C.,And Potter,S.(2013). 보존 정책 계획에서 진화론 적 관점의 중요성. 몰. 에콜. 22, 5969–5971. 도이:10.1111/멕.12565
PubMed 추상|CrossRef 전체 텍스트|Google 학술검색
모리츠,C.,프랫,R.C.,은행,S.,Bourke,G.,브래그,J.G.,대담한,P.,et al. (2018). 호주 도마뱀(Gehyra)의 종 복합체에서의 비밀 혈통 다양성,신체 크기 발산 및 sympatry. 진화 27,54-66. 도이:10.1111/에보.쩔짤쨌짱쨘째쩍 짹쨍쨌짹쨍 쨉챨철쨌 32 짹챈 30,13 첸 쩔짙쩔징징쩔짙째챠쨉쨉()째챠째쨈쩌쩐(1688-7662 (2009). 보전에 대한 분류 학적 변화의 영향: 그것은 죽입니까,저장할 수 있습니까,아니면 그냥 관련이 없습니까? 바이올. 단점. 142, 3201–3206. doi:10.1016/j.biocon.쩔짤쨌짱쨘째쩍 짹쨍쨌짹쨍 쨉챨철쨌 32 짹챈 30,13 첸 쩔짙쩔징징쩔짙째챠쨉쨉()째챠째쨈쩌쩐(1688-7662 호주 사막의 봄 amphipods 에서 특별한 마이크로 endemism. 몰. 계통 발생. 에볼. 66, 645–653. 도이:10.1016/j.ympev.2012.10.013
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Murphy,N.P.,King,R.A. 및 Delean,S.(2015). 종,ESUs 또는 개체군? 호주 사막의 봄 amphipods 에서 형태 학적으로 비밀스러운 다양성을 구분하고 설명합니다. 반전. 시스트. 29, 457–467. doi:10.1071/IS14036
CrossRef 전체 텍스트|Google Scholar
Nater,A.,Mattle-Greminger,M.P.,Nurcahyo,A.,Nowak,M.G.,de Manual,M.,Desai,T.,et al. (2017). 새로운 오랑우탄 종에 대한 형태 측정,행동 및 게놈 증거. 커르. 셀 바이올. 27, 3487–3498. 도이:10.1016/j.새끼.쩔짭짹쨍짱쨘째쩍 짹쨍쨌짹쨍 쨉챨철쨌 32 짹챈 30,13 첸 쩔짙쩔징징쩔짙째챠쨉쨉()째챠째쨈쩌쩐(1688-7662,외. (2016). 호주 Pelargonium 에서 genotyping-by-sequencing 을 통한 인구 및 계통 발생 분해. 몰. 에콜. 25, 2000–2014. 도이:10.1111/멕.13584
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Nistelberger,H.,Byrne,M.,Coates,D.,And Roberts,J.D.(2014a). 노래기에서 강한 계통 학적 구조는 반 건조 호주에서 줄무늬 철 형성에 대한 개체군 간의 홍적세 대변 성을 나타냅니다. PLoS 하나 9:e0093038. doi:10.1371/저널.폰.0093038
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Nistelberger,H.,Gibson,N.,Macdonald,B.,Tapper,S.-L.,and Byrne,M.(2014b). 생물 다양성 핫스팟에서 두 개의 mesic refugia 에 대한 계통 학적 증거. 유전 113,454-463. doi:10.1038/hdy.쩔짤쨌짱쨘째쩍 짹쨍쨌짹쨍 쨉챨철쨌 32 짹챈 30,13 첸 쩔짙쩔징징쩔짙째챠쨉쨉()째챠째쨈쩌쩐(1688-7662 짐승의 계산 성능 및 통계적 정확성 및 다른 방법과의 비교. 시스트. 바이올. 65, 381–396. 도이:10.1093/sysbio/syv118
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Oliver,P.M.,Keogh,J.S. 및 Moritx,C.(2015). 진화론 적 다양성을 목록 화하고 이해하는 새로운 접근법:호주 헤르페톨로지의 관점. 오스트. 제이 졸. 62, 417–430. doi:10.1071/ZO14091
CrossRef 전체 텍스트|Google Scholar
Oliver,P.M.,Laver,R.J.,Demello Martins,F.,Pratt,R.C.,Hunjan,S.,And Moritz,C.(2017). 열대성 호주의 척추 동물 종말론과 진화론 적 폐허의 새로운 핫스팟. 다이버. 디스트립. 23, 53–66. doi:10.1111/ddi.쨈챘짹쨀째쩔징 쨉청쨋처 쨘쨍쩔징 쨈챘쩔짤 쨘쨍쩔징 쨈챘쩔짤 쨘쨍쩔징 쨈챘쩔짤 쨘쨍쩔징 쨈챘쩔짤 쨘쨍쨈쨈. 식물 및 동물학 코드는 이름없는 새로운 종의 출판을 낙담시킴으로써 생물 다양성 연구를 방해합니다. 분류군 59,1201-1205. 쨈챘짹쨀째쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔징 쨈챘쩔짤 쨘쨍쨀쨩쨈쨈. 분류학의 통합적인 미래. 앞. 주울. 7:16. 도이:10.1186/1742-9994-7-16
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Palsbøll,P.J.,Bérubé,M.,and Allendorf,F.W.(2007). 인구 유전 데이터를 사용하여 관리 단위의 식별. 트렌드 에콜. 에볼. 22, 11–16. 도이:10.1016/j.나무.2006.09.003
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Patton,Jl,And Smith,M.F.(1994). Pocket Gophers(속 Thomomys)에서 Paraphyly,polyphyly 및 종 경계의 성격. 시스트. 바이올. 43, 11–26. doi:10.1093/sysbio/43.1.쩔짤쨌짱쨘째쩍 짹쨍쨌짹쨍 쨉챨철쨌 32 짹챈 30,13 첸 쩔짙쩔징징쩔짙째챠쨉쨉()째챠째쨈쩌쩐(1688-7662 (2016). 3 개 대륙에서 위협받는 종 복구 계획의 유전 적 요인. 앞. 에콜. 에비론. 14, 433–440. doi:10.1002/요금.1323
CrossRef 전체 텍스트|Google Scholar
Potter,S.,Bragg,J.G.,Peter,B.M.,Bi,K.,And Moritz,C.(2016). 팁의 계통 발생학:열대 도마뱀 인 Carlia amax 에서 계통과 인구 통계 학적 역사를 유추합니다. 몰. 에콜. 25, 1367–1380. 도이:10.1111/멕.13546
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Potter,S.,Xue,A.T.,Bragg,J.G.,Rosauer,D.F.,Roycroft,E.J.,And Moritz,C.(2018). 홍적세의 기후 변화는 열대 사바나 전역의 다양 화를 유도합니다. 몰. 에콜. 27, 520–532. 도이:10.1111/멕.14441/full
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Rabosky,D.L.,Hutchinson,M.N.,Donnellan,S.C.,Talaba,A.L.,And Lovette,I.j.(2014). 호주 skinks(Scincidae:Ctenotus)의 광범위한 그룹에서 종 경계의 계통 발생 분해. 몰. 계통 발생. 에볼. 77, 71–82. 도이:10.1016/j.ympev.2014.03.026
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Ralls,K.,Ballou,J.D.,Dudash,M.R.,Eldridge,M.D.B.,Fenster,C.B.,Lacy,R.C.,et al. (2018). 분열 된 개체군의 유전 적 관리에서 패러다임 변화를 요구한다. 콘서브. 레트. 11, 1–6. 도이:10.1111/conl.124126
CrossRef 전체 텍스트|Google Scholar
Rannala,B.(2015). 종 구분의 예술과 과학. 커르. 주울. 61, 846–853. doi:10.1093/czoolo/61.5.846
CrossRef 전체 텍스트|Google 학술검색
리델,A.,Sagata,K.,Suhardjono,Y.R.,Tänzler,R.,그리고 Balke,M.(2013). 패스트 트랙에서의 통합 분류 체계-생물 다양성 연구에서 더 많은 지속 가능성을 향해. 앞. 주울. 10:15. doi:10.1186/1742-9994-10-15
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Rieseberg,L.H.,And Willis,J.H.(2007). 식물 종화. 과학 317,910-914. 도이:10.1126/과학.1137729
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Rosauer,D.F.,Blom,M.P.K.,Bourke,G.,Catalano,S.,Donnellan,S.,Mulder,E.,et al. (2016). 계통 지리학,핫스팟 및 보존 우선 순위:호주 최고급 사례. 바이올. 단점. 204, 83–93. doi:10.1016/j.biocon.2016.002
CrossRef 전체 텍스트|Google 학술검색
Rosauer,D.F.,학적,M.,Blom,M.P.K.,Coates,D.J.,데 클란 도넬란,S.,대담한,P.,et al. (2018). 호주 킴벌리(Kimberley)의 진화 적 다양성을위한 실제 보존 계획은 불확실한 분류법 Conserv 를 회피합니다. 레트. 11:e12438. 도이:10.1111/conl.12438
CrossRef 전체 텍스트|Google 학술검색
베,E.B.,사버,B.A.J.,갈색,J.W.,Des Roches,S.,하드윅,K.M.,Hether,D.T.,et al. (2012). Goldilocks 는 Santa Rosalia 를 만납니다:임시 분화 모델은 시간 척도에 걸쳐 다양 화 패턴을 설명합니다. 에볼. 바이올. 39, 255–261. 도이:10.1007/s11692-012-9171-x
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Roux,C.,Fraïsse,C.,Romiguier,J.,Anciaux,Y.,Galtier,N. 및 Bierne,N.(2016). 게놈 발산의 연속체를 따라 분화의 회색 영역에 빛을 흘리기. 플로스 바이올. 14:e2000234. doi:10.1371/저널.pbio.2000234
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Ryder,O.A.(1986). 종 보존 및 계통 학-아종의 딜레마. 트렌드 에콜. 에볼. 1, 9–10. 도이:10.1016/0169-5347(86)90059-5
CrossRef 전체 텍스트/Google Scholar
Sampson,J.F.,Byrne,M.,Yates,C.J.,Gibson,N.,Thavornkanlapachai,R.,Stankowski,S.,et al. (2014). 현대의 꽃가루 매개 유전자 이민은 조각난 풍경 속에서 희귀 한 관목의 역사적 고립을 반영합니다. 유전 112,172-181. doi:10.1038/hdy.2013.89
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Santamaría,L.,and Méndez,P.f.(2012). 생물 다양성 정책의 진화:현재의 격차와 미래의 요구. 에볼. 아플. 5, 202–218. 도이:10.1111/j.1752-4571.2011.00229.x
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Schlick-Steiner,B.C.,Steiner,F.M.,Seifert,B.,Stauffer,C.,Christian,E.,and Crozier,R.h.(2010). 통합 분류학:생물 다양성을 탐구하기위한 다중 소스 접근법. 아누. 목사 엔토몰. 55, 421–438. doi:10.1146/annurev-ento-112408-085432
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Seehausen,O,Takimoto,G.,Roy,D.,And Jokela,J.(2008). 변화하는 환경에서 하이브리드 화를 통한 종화 역전 및 생물 다양성 역학. 몰 에콜. 17, 30–44. 도이:10.1111/j.1365-294X.2007.03529.x
PubMed 추상|CrossRef 전체 텍스트|Google 학술검색
목자,K.A.,Thiele,K.R.,Sampson,J.,Coates,D.J.,그리고 학적,M.(2015). 희귀,새로운 종의 Atriplex(명아주과)구성하는 두 개의 유전한 암호 형태학에서 인구조 웨스턴 오스트레일리아:에 대한 영향 분류 및 보존. 오스트. 시스트. 봇. 28, 234–245. 도이:10.1071/SB15029
CrossRef 전체 텍스트|Google Scholar
Singhal,S. 및 Bi,K.(2017). 역사는 엉망진창을 정리합니다:열대성 봉합 구역에서의 발산 및 내향 운전에서의 시간의 영향. 진화 71,1888-1899. 도이:10.1111/에보.13278
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Singhal,S.,Hoskin,C.,Couper,P.,Potter,S.,And Moritz,C.(2018). 비밀 종을 해결하기위한 프레임 워크:호주 젖은 열대 지방의 도마뱀에서 사례 연구. 시스트. 바이올. . 도이:10.1093/sysbio/syy
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Singhal,S.,And Moritz,C.(2013). 계통 학적 계통 사이의 생식 격리는 발산으로 확장됩니다. 프록. 로이. Soc B280:20132246. doi:10.1098/rspb.2013.2246026
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Steane,D.A.,Potts,B.M.,McLean,E.,Collins,L.,Prober,S.M.,Stock,W.D.,et al. (2015). 게놈 전체 스캔은 건조한 적응 유칼립투스에서 비밀스러운 인구 구조를 나타냅니다. 트리 제네. 게놈 11:33. 도이:10.1007/s11295-015-0864-쨩챌쩐첨쨉챤쨌쨔첩짙:102-81-571-57|챘쩍쨍쩐첨쩍째챠:짝 2014-쩌짯쩔챦짱쨘째쩍 짹쨍쨌짹쨍 쨉챨철쨌 32 짹챈 30,13 첸 쩔짙쩔징징쩔 (2017). 비밀 종에 숨겨진 진화 과정을 찾는 것. 트렌드 에콜. 에볼. 33, 153–163. 도이:10.1016/j.나무.2017.11.007
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Sukumaran,J.,And Knowles,L.L.(2017). 다종 합체는 종족이 아닌 구조를 구분합니다. 프록. 냇. 아카. Sci. 미국 114,1607-1612. doi:10.1073/pnas.1607921114
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Swarts,N.D.,Clements,M.A.,Bower,C.C.,And Miller,j.t.(2014). 형태 학적으로 유사하고 성적으로기만적이고 고도로 멸종 위기에 처한 난초의 복합체에서 보존 단위를 정의합니다. 바이올. 단점. 174, 55–64. doi:10.1016/j.biocon.쨩챌쨋첨쨉챤쨌쨔첩짙:506-81-571-57 챘쩍쨍쩐첨쩍째챠:짝 2014.03.017
CrossRef Full Text|Google Scholar
Tautz,D.,Arctander,P.,Minelli,A.,Thomas,R.H.,And Vogler,A.P.(2003). DNA 분류법에 대한 항변. 트렌드 에콜. 에볼. 18, 70–74. 도이:10.1016/S0169-5347(02)00041-1
CrossRef 전체 텍스트|Google Scholar
Taylor,H.R.,Dussex,N. 및 van Heezik,Y.(2017). 보존 실무자에 대한 이행 장벽을 확인함으로써 보존 유전학 격차를 해소한다. 글롭. 에콜. 콘서브. 10, 231–242. doi:10.1016/j.gecco.2017.04.001
CrossRef 전체 텍스트|Google Scholar
Thomson,S.A.,Pyle,R.L.,Ahyong,S.T.,Alonso-Zarazaga,M.,Ammirati,J.,Araya,J.F.,et al. (2018). 과학에 기반한 분류학은 세계 보전에 필요합니다. 플로스 바이올. 16:e2005075. 도이:10.1371/저널.pbio.2005075
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Vieites,D.R.,Wollenberg,K.C.,Andreonec,F.,Köhler,J.,Glaw,F.,and Vences,M.(2009). 마다가스카르의 생물 다양성에 대한 광대 한 과소 평가는 통합 양서류 재고에 의해 입증되었습니다. 프록. 나틀. 아카드. Sci. 미국 106,8267-8272. doi:10.1073/pnas.나는 이것이 내가 할 수있는 유일한 방법이라고 생각한다. 보존 관리 단위 진단. 단점. 바이올. 8, 354–363. 도이:10.1046/j.1523-1739.1994.08020354.나는 이것이 내가 할 수있는 유일한 방법이라고 생각한다. 태평양 연어,Oncorhynchus spp.,그리고 멸종 위기에 처한 종의 법에 따른”종”의 정의. 마르. 물고기. 목사 53,11-22.쨈챘짹쨀째쩔징 쨉청쨋처 쨘쨍쩔징 쨈챘쩔짤 쨘쨍쩔징 쨈챘쩔짤 쨘쨍쨈쨈. “진화론적으로 중요한 단위와 생물다양성 보전에는 멸종 위기에 처한 종 행위”에서 발전과 수중 생태계:정의한 단위에서 인구의 보존,eds J.L.Nielsen(베데스다,메릴랜드:미국의 어업 사회),8-27.
Google 학술검색
웨이플스,R.S.,구스타프손,R.G.,Weitkamp,L.A.,Myers,J.M.,Jjonson,O.W.,버스바,P.J.,et al. (2001). 태평양 북서부의 연어의 다양성을 특징으로합니다. 제이 피쉬 바이올. 59, 1–41. 도이:10.1111/j.1095-8649.2001.tb01376.x
CrossRef 전체 텍스트|Google 학술검색
웨이플스,R.S.,Nammack,M.,코크,J.F.,및 허칭,J.A.(2013). 두 가지 행위에 대한 이야기:캐나다와 미국의 멸종 위기 종 목록 관행. 생물 과학 63,723-734. 도이:10.1093/생명 과학/63.9.723
CrossRef 전체 텍스트|Google Scholar
Waples,R.S.,Punt,A.E.,And Cope,J.m.(2008). 해양 자원 관리에 유전자 데이터 통합:우리는 어떻게 더 잘 할 수 있습니까? 물고기 9,423-449. 도이:10.1111/j.1467-2979.2008.00303.쨈챘짹쨀째쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔챘짹쨀째쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔챘짹쨀째쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔챘짹쨀째쩔징 쨈챘쩔짤 쨘쨍쨀쨩쩔챘짹쨀째쩔징 쨈챘쩔짤 쨘쨍쨈쨈. 생물 다양성 경관에서의 전략적 분류법:희귀하고 잘 알려지지 않은 식물 군에 대한 보전 결과를 극대화하기위한 새로운 접근법. 생물 생물. 콘서브. 24, 17–32. doi:10.1007/s10531-014-0785-4
CrossRef 전체 텍스트|Google Scholar
Whitlock,R.(2014). 적응적이고 중립적 인 유전 적 다양성과 생태 학적 구조와 기능 사이의 관계:메타 분석. 제이 에콜. 102, 857–872. doi:10.1111/1365-2745.12240
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Wiley,E.O.(1978). 진화 종의 개념이 재검토되었다. 시스트. 바이올. 27, 17–26. doi:10.2307/2412809
CrossRef 전체 텍스트|Google Scholar
Wilson,E.O.(2017). 생물 다양성 연구는 지상에 더 많은 부츠를 필요로합니다. 냇. 에콜. 에볼. 1, 1590–1591. doi:10.1038/s41559-017-0360-y
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Yang,Z.H. 및 Rannala,B.(2014). 여러 loci 의 DNA 서열 데이터를 사용하여 유도되지 않은 종 구분. 몰. 바이올. 에볼. 31, 3125–3135. doi:10.1093/molbev/msu279
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Yeates,D.K.,Seago,A.,Nelson,L.,Cameron,S.L.,Joseph,L.,And Trueman,W.h.(2011). 통합 분류법 또는 반복 분류법? 시스. Ent. 36, 209–217. doi: 10.1111/j.1365-3113.2010.00558.x
CrossRef Full Text | Google Scholar