i början av XIX-talet observerade von Reckingausen att blodkärlen täcktes av ett cellark. Detta encells tjocka skikt består av endotelceller som leder den inre ytan av blod och lymfkärl. Hos människor uppskattas att den totala endotelskiktsytan är cirka 35 m2 och att det finns cirka 1 till 1,6 103 endotelceller. Den totala längden på artärer, vener och kapillärer är cirka 90000 km (mer än att resa två gånger runt om i världen). Endotelceller är plana och förbundna mellan varandra genom cellkorsningar. Deras huvudsakliga funktion är som mellanhänder mellan blodet och de andra vävnaderna och bidrar till blodegenskaper, både i normala och patologiska vävnader. De är också viktiga för utbyte av immunceller mellan blod och vävnader.
1. Morfologi
endotelceller är mycket platta celler, så mycket att deras kärna är cellens högre struktur, även om kärnan också är plattad. Den cellulära formen är anpassad till kanalen endotelcellen är foder (Figur 1, 2 och 3). I de smalare kapillärerna kan endotelceller förlänga sin cytoplasma hela blodkärlets omkrets, så att kanalen är en rad endotelceller. I större kanaler, som artärer och vener, behövs många endotelceller för att täcka kanalens totala omkrets.
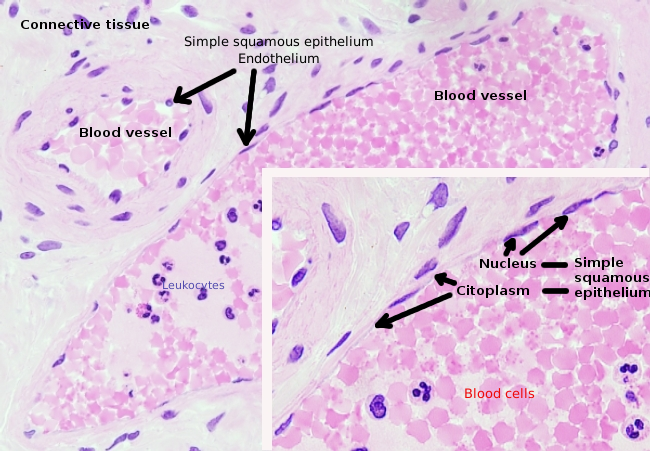
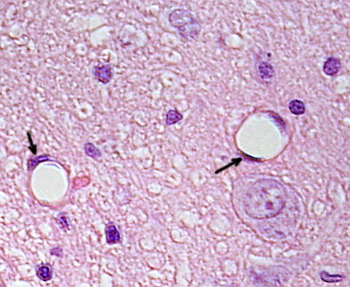
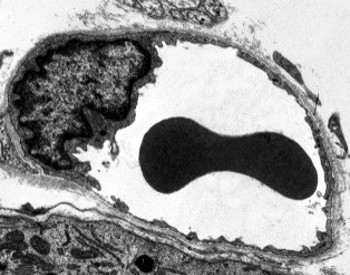
Perinukleär cytoplasma innehåller de flesta mitokondrier och andra organeller, såsom Golgi-apparater, medan perifercytoplasma är tunnare och har få organeller, även om endoplasmatisk retikulum kan hittas nära plasmamembranet. Endotelceller har många vesiklar, de flesta är endocytiska vesiklar.
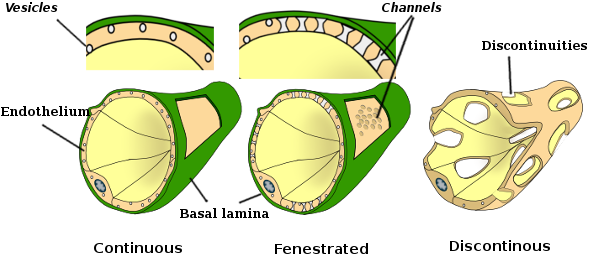
i vissa regioner, såsom lever, njurbark och endokrina körtlar, visar endotelceller i kapillärer porer eller små passager, kända som fenestratrioner. De är vägar på cirka 60 till 70 nm i diameter, med en smal passage på cirka 5 nm, som direkt kommunicerar blodet och lymfen med de andra vävnaderna. Kapillärer som har dessa typer av endotelceller är kända som fenestrerade kapillärer. Porer fördelas i grupper och deras densitet beror på typen av endotel.
endotelceller separeras från den omgivande vävnaden med ett lager av extracellulär matris som kallas basal lamina. Endotelcell bidrar till detta skikt med proteinerna laminin, fibronektin och kollagen typ II, IV och V.
källa och proliferation
Vaskulogenes är bildandet av nya endotelceller i vävnader utan redan existerande blodkärl. Det händer i embryot. Hos vuxna, och även i embryon, genereras nya blodkärl, och därför nya endotelceller, från redan närvarande blodkärl. Denna process kallas angiogenes. Nya endotelceller produceras genom förgrening eller delning av blod och lymfkärl. Angiogenes är vanligt i växande vävnader eller de under tung ombyggnad, som livmodern hos däggdjurshonor, eller i patologier som tumörer, inflammation och sår.
Vaskulogenes
första endotelceller hos däggdjur genereras efter gastrulation under embryonutvecklingen. De skiljer sig från celler som kallas angioblaster, bildar grupper och organiseras i korta blodkärl. Denna process sker först i embryonets vitelinsäck. Dessa initiala blodkärl växer och ansluter mellan varandra för att bilda ett nätverk. Senare rekryterar de fibroblaster och muskelceller. Detta initiala nätverk genomgår en kontinuerlig ombyggnadsprocess under nästa utvecklingsstadium. I embryon är alla endotelceller initialt lika, men de differentieras senare i endotel i ven, artär, kapillär och lymfkärl. Flera molekyler som FGF2, BMP4, IHH och VEGF inducerar separerade differentieringsvägar. Notch inducerare verkar vara mycket viktigt eftersom det leder till artärendotel, medan dess hämning leder till venendotel. Lymfatiskt endotel bildas från kardinalvenendotelet och behöver uttrycket av COUPF-II och SOX18. Dessutom differentieras endotelet i enlighet med det organ det är beläget i. Till exempel bildar endotel ett tätt förseglat skikt i hjärnan för att bilda blod-hjärnbarriären, medan det fenestreras av löst organiserat i levern för att gynna utbytet av molekyler med blodet.
eftersom endotelceller organiseras i artär-och venkanaler ger vissa endotelceller upphov till hematopoietiska celler genom en differentieringsprocess som kallas endotel-hematopoietisk övergång (EHT). Denna process händer också i vissa vuxna vävnader som under bildandet av placenta och andra vävnader under graviditeten.
angiogenes
angiogenes är bildandet av nya blodkärl och endotel genom förgrening eller delning av redan existerande blodkärl. Hos vuxna är proliferationshastigheten för endotelceller mycket låg. Till exempel hos däggdjur kan en endotelcell dela sig en gång per månad eller vänta flera år. Det betyder att angiogenes är sällsynt i vävnader under normal fysiologi, förutom kvinnliga reproduktionsorgan. Det kan dock aktiveras i patologiska processer som vävnadsskador och tumörer. Vissa ämnen inducerar angiogenes, såsom endoteltillväxtfaktor (EGF), sur EGF, fibroblasttillväxtfaktor (FGF), transformerande tillväxtfaktor (TGF) och prostaglandiner. Angiogenes börjar när angiogena ämnen frigörs från omgivande vävnader. Endotelcellen i ett nära blodkärl, vanligtvis från en venule, börjar migrera till den angiogena signalkällan och drar de andra endotelcellerna, som börjar proliferera och bilda ett nytt kärl. Dessa endotelceller förlorar tillfälligt sina täta anslutningar vilket resulterar i avtäckning av basalmembranet, vilket snabbt smälts av enzymer som frigörs från endotelcellerna. Vissa endotelceller går igenom det smälta basalmembranet medan spridningen fortsätter. All denna process möjliggör förlängning och förgrening av det nya blodkärlet tills vaskulogenessignalen försvinner. Spetsarna på blodkärlen smälter samman med andra blodkärl för att bilda slutna kretsar och möjliggör flöde av blod utan döda ändar.
regioner med endotelcellsförfäder har hittats i dorsal aorta och endokardium. Mesenkymala celler kan differentieras från endotelceller i endokardiet och bilda tricuspidventilen och vissa fibroblaster i hjärtat, men inte kardiomiocyter. Endotelceller är så förnuftiga för externa signaler att hålla endotelintegriteten är en aktiv process, som förmedlas av andra signaler som FGF. Den aktiva processen är inte bara för integritet utan för att behålla den cellulära fenotypen också. När endotelceller inte får korrekta signaler dör de av apoptos eller blir mesenkymala celler som kan syntetisera riklig extracellulär matris. Övergången från endotel till mesenkymcell ligger bakom vissa sjukdomar som arterioskleros och fibrös miokardit.
funktion
tanken att endotelet bara är ett passivt foderskikt av lymfatiska och blodkärl måste ändras. Endotelfunktioner är varierade och väsentliga för organismen. Det är faktiskt en fysisk struktur av kardiovaskulära och lymfatiska kanaler, men det reglerar också utbytet av molekyler mellan kärlets lumen och de omgivande vävnaderna, och det påverkar blodets fysiologiska egenskaper. Förutom en fysisk barriär utvecklar endotelceller sekretoriska, metaboliska och immunfunktioner. De förändrar sitt fysiologiska beteende som påverkas av molekyler som tillväxtfaktorer, koagulanter och antikoagulantia, lågdensitetslipoproteiner, kväveoxid, serotonin, enkefalin och många andra. Endotelceller har receptorer för alla dessa ämnen.
barriär
endotelceller bildar ett skikt som vanligtvis fungerar som en barriär mellan blodet och de omgivande vävnaderna. Sammanhållningen mellan endotelceller uppfylls av cellkorsningar, såsom täta korsningar och vidhäftande korsningar. Gapkorsningar har också observerats, även om deras huvudsakliga funktion är kommunikationen mellan angränsande celler. Endotelceller kan modulera dessa vidhäftningar och ändra barriärens permeabilitet, vilket kan påverka inte bara molekyler utan också celler som passerar endotelet. Men i vissa organ som lever är endotelceller mycket löst förpackade och lämnar mycket ledigt utrymme så att endotel knappast kan betraktas som en barriär.
Överföringselektronmikroskopibilder visar många vesiklar i cytoplasman hos endotelceller. De tros vara involverade i intracellulär transport mellan apikalen (mot blodet) och basolaterala membran (mot basala lamina). Denna typ av transport är känd som transcytos. Det är intressant att transcytosblåsorna är mer rikliga i kapillärernas endotelceller än i större blodkärl. Det indikerar att endotelcellerna i kapillärerna har en mer intensiv utbyte av molekyler mellan blod och vävnader, medan de i kärl med stor diameter huvudsakligen är involverade i att leda blodet. Vissa endotelceller, kända som fenestrerade, har porer eller mycket små kanaler som direkt förbinder blodet med de omgivande vävnaderna, vilket gör att vissa små molekyler kan korsa endotelet utan att komma in i cytoplasman hos några endotelceller (Figur 4). Slutligen finns det organ med mycket permeabelt endotel. I levern är sinusoider blodkärl där endotelceller lämnar ledigt utrymme mellan varandra, och en funktion som barriär finns knappast.
många celltyper reser i blodet mot sina målorgan i kroppen. Där korsar de blodkärlens endotel, vanligtvis i nivå med postkapillära vener. Det betyder att endotelceller måste modifiera cellkorsningarna för att låta celler gå igenom endotelskiktet. Leukocyter lämnar blodkärlet genom att känna igen och förankra till specifika molekyler i det apikala membranet i endotelcellerna. Selektiner, integriner och immunoglobuliner är ansvariga för igenkänning och vidhäftning av leukocyter till endotelskiktet. Selektiner börjar förankring av leukocyter, som rullar över endotelytan. Denna initiala vidhäftning är svag och reversibel.
många leukocyter lämnar blodet under inflammatoriska processer för att flytta till de drabbade vävnaderna. Kemokiner är leukocyt-attraktiva molekyler som frigörs av skadade vävnader och kopplas till glykokalix av endotelceller. Leukocyter som rullar över endotelytan med kemokiner aktiveras, cellcelladhesion blir starkare och leukocyt förblir på plats. Aktiveringen av leukocyten leder till aktiveringen av dess integriner, som känner igen endotelcellens immunoglobuliner. Dessa immunoglobuliner uttrycks i endotelcellmembranen efter aktiveringen av cellen med kemokiner. Cell-celladhesion höjer kalciumkoncentrationen i endotelceller och leder till cellkorsningar disorganisering och cytoplasmindragning. På detta sätt kan leukocyter flytta till gränsen för endotelceller och korsa endotelet. Adhesionsmolekyler är också involverade i denna rörelse av leukocyter.
blodegenskaper
endotelfunktionerna är mer komplexa än att bara kontrollera molekylerna och cellerna som går över endotelet. Endotelceller är också involverade i blodtryck, koagulering och några andra blodegenskaper.
primitivt cirkulationssystem tros dyka upp för 600 miljoner år sedan hos ryggradslösa djur, men det saknade endotel. Endotelceller uppträdde för 100 miljoner år sedan och gav blod ett mer laminärt flöde (inte turbulent) och därför en effektivare gasutbyte.
endotelceller modulerar blodtrycket genom att frigöra ämnen som verkar på blodkärlens glatta muskel. De släpper ut kväveoxid (NO) och prostacyklin, som slappnar av vaskulär glatt muskel. De släpper också endotelin och blodplättaktivatorfaktorn, båda minskar blodkärlets diameter. Kväveoxid frigörs konstitutivt och ger en korrekt muskelton, hämmar trombocytaggregation och leukocytadhesion. Endotelin är en stark vasokonstriktor. Hur endotelceller bestämmer vilken molekyl som ska släppas är ännu inte klart, men mekanorreceptorer i deras apikala membran som kan känna blodets flödesegenskaper kan vara troliga.
under normala förhållanden släpper endotelceller molekyler i blodet som hjälper till att upprätthålla en korrekt fluiditet. De arbetar på två nivåer: fluiditet (antikoagulantia) och förhindrar trombocytaggregation (antitrombotika). Proteiner C och S är viktiga molekyler som påverkar blodfluiditeten. C-protein, som bildar ett komplex med S-protein, inaktiverar koagulationsfaktorer VIIIa och Va. S-protein syntetiseras av endotelceller. Vidare innehåller endotelglykokalixen en glykosaminoglykan som liknar heparin som kan inaktivera trombin.
relaterat till deras antikoagulantfunktion frigör endotelceller kväveoxid och prostacyklin, båda stiger den cykliska AMP i blodplättar och gör svårare trombocytaggregation. Dessa två molekyler släpps kontinuerligt ut i blodet. Endotelceller har ektonukleaser i sina apikala membran, som avlägsnar ATP och ADP, båda starka promotorer av trombocytaggregation. Dessutom frigör endotelceller en aktivatormolekyl som omvandlar plaminogen till plasmin, vilket gynnar avlägsnande av trombi.
alla dessa molekylära vägar kan förändras när endotelceller får vissa signaler eller vävnad skadas, vilket leder till blodkoagulering och trombocytaggregation. Under dessa omständigheter blir endotelceller sedan aktiva deltagare i koagulering och trombos.
immunförsvar
endotelceller spelar en viktig roll i immunförsvaret och är involverade i två mekanismer: antigenpresentation till T-lymfocyter och rekrytering av immunceller. Tillsammans med makrofager kan endotelceller presentera antigener till T-lymfocyter eftersom de konstitutivt uttrycker MHC-i (major histokompatibilitetskomplex) och kan induceras för att uttrycka MHC-II, båda nödvändiga för antigenpresentation. Endotelceller kan aktivera immunminne, men inte nya T-lymfocyter. Det finns dubbelriktad aktivering mellan endotelcell och T-lymfocyter, så att endotelcellsfrisättningsmolekyler för att locka inflammatoriska celler och uttryckliga vidhäftningsmolekyler för förankring av blodleukocyter.
bibliografi
Cines BD, Pollak ES, Buck CA, Loscalzo J, Zimmerman GA, McEver RP, Pober JS, Wick TM, Konkle BA, Schwartz BS, Barnathan ES, McCrae KR, kram BA, Schmidt A-M, Stern DM 1998. Endotelceller i fysiologi och i patofysiologiav kärlsjukdomar. Tidningen för american society of hematology. 91:3527-3561 ![]()
Fajardo LF. 1988. Komplexiteten hos endotelceller. American journal of clinical pathology. 92:241-250.
Michiels C. 2003. Endotelcellsfunktioner. Journal of cellular physiology. 196: 430-443 ![]()