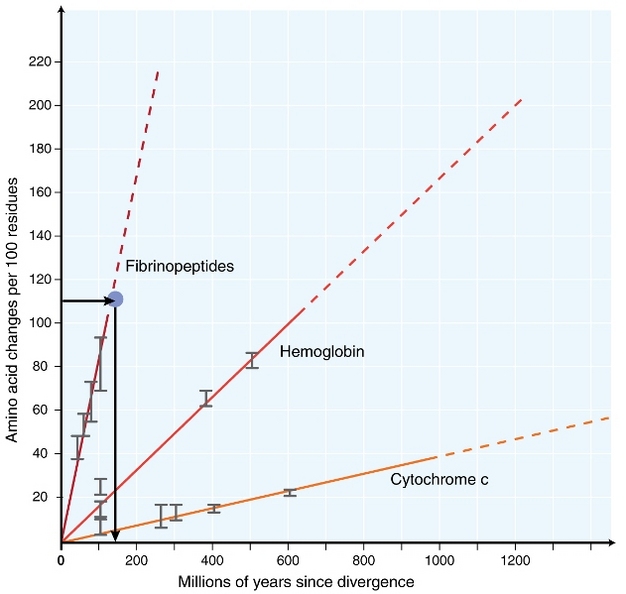
In het begin van de jaren 1960, biologen begonnen te onderzoeken hoe eiwitten in verschillende soorten evolueren in de volgorde van niveau (Zuckerkandl 1962, Margoliash 1963, Doolittle & Blombäck 1964). De geanalyseerde eiwitten omvatten hemoglobine (Zuckerkandl & Pauling 1962), cytochroom C (Margoliash 1963) en fibrinopeptiden (Doolittle & Blombäck 1964). Deze vroege onderzoeken leidden tot een opmerkelijke ontdekking: het bleek dat het aantal verschillen tussen eiwitsequenties van verschillende soorten ongeveer evenredig was met de tijd sinds de divergentie van soorten (figuur 1).
Zuckerkandl&Pauling (1965) vergeleek de constante accumulatie van aminozuursubstituties in de loop van de tijd met regelmatige ’teken’ van klokken, en stelde dat ‘er een moleculaire evolutionaire klok kan bestaan’. Aldus, werd de term ‘moleculaire klok’ aanvankelijk bedacht om veranderingen in aminozuren die in proportioneel aan tijd sinds species divergentie voorkomen te beschrijven.
sinds het eerste gebruik wordt de term ‘moleculaire klok’ in veel verschillende contexten gebruikt. Tegenwoordig wordt het vaak gebruikt om simpelweg te verwijzen naar het aantal veranderingen, of ‘substituties’, geaccumuleerd in de opeenvolging van DNA of eiwitten in een bepaalde afstamming. Het aantal substituties per een bepaalde tijdseenheid kan worden omschreven als de’ snelheid ‘van de moleculaire klok, die in deze context gelijk is aan’evolutionaire snelheid’. Merk op dat de oorspronkelijke connotatie in sommige gevallen nog steeds geldt. Bijvoorbeeld, wanneer mensen zeggen ‘we veronderstelden een moleculaire klok’ (met name in fylogenetische analyses), betekent dit dat het aantal substituties werd verondersteld om constant te voorkomen in de tijd.
het concept van een constante moleculaire klok heeft buitengewone implicaties voor evolutionaire biologen. Als een constante moleculaire klok zoals aanvankelijk voorgesteld echt bestond, zou het afleiden van de timing van evolutionaire gebeurtenissen een vrij eenvoudig probleem worden (figuur 1). Het is echter overduidelijk geworden dat substituties niet constant in de loop van de tijd voorkomen in verschillende geslachten (Kumar 2005). Niettemin is het concept van moleculaire klok zeer invloedrijk geweest op het gebied van moleculaire evolutie. Een van de belangrijkste ideeën geïnspireerd door het concept van de moleculaire klok is de neutrale theorie van moleculaire evolutie.
de moleculaire klok en neutraliteit
toen het idee van een constante moleculaire klok voor het eerst naar voren kwam, werd gedacht dat de belangrijkste evolutionaire kracht die ten grondslag ligt aan aminozuur-of nucleotidesubstituties natuurlijke selectie was. Volgens deze denkwijze zou een constante moleculaire klok erop wijzen dat adaptieve substituties in verschillende soorten constant in de tijd voorkomen. Het is echter moeilijk uit te leggen hoe adaptieve substituties zich op zo ‘ n klokachtige manier zouden voordoen. Theoretisch wordt het lot van adaptieve mutaties bepaald door verschillende evolutionaire parameters, zoals de sterkte van het selectieve voordeel van die mutatie, de grootte van de effectieve populatie en de adaptieve mutatiesnelheden (Kimura 1983). Deze parameters zullen waarschijnlijk verschillen tussen species, en zelfs binnen een species, afhankelijk van specifieke veranderingen en hun interactie met milieu ‘ s. in plaats daarvan stelde Kimura (1968, 1969) voor dat de meeste veranderingen op moleculair niveau weinig functionele gevolgen hebben of ‘neutraal’zijn. Als een mutatie geen fitness-gevolg heeft, wordt het lot in de populatie volledig bepaald door willekeurig toeval. Dit betekent dat we niet kunnen voorspellen of een specifieke neutrale mutatie uiteindelijk in de populatie zal worden gefixeerd. Echter, de snelheid waarmee neutrale substituties optreden in de populatie kan worden voorspeld omdat het afhangt van een enkele parameter, namelijk de mutatiesnelheid (Kimura 1968).
stel je een populatie voor met N aantal haploïde individuen. Als neutrale mutaties optreden met een snelheid u per individu per generatie, zal het totale aantal mutaties in één generatie N keer u zijn. omdat al deze nieuwe mutaties neutraal zijn, wordt hun lot volledig bepaald door toeval. Met andere woorden, alle mutaties hebben gelijke kans op het bereiken van fixatie (wat leidt tot een ‘substitutie’). De kans dat elke nieuwe neutrale mutatie fixatie zal bereiken, gegeven het feit dat een substitutie heeft plaatsgevonden, is gewoon 1 / N. Het percentage substituties wordt berekend als het aantal nieuwe mutaties in elke generatie (Nu) vermenigvuldigd met de kans dat elke nieuwe mutatie fixatie bereikt (1 / N), die gelijk is aan u. met andere woorden, voor neutrale mutaties is de substitutiesnelheid gelijk aan de mutatiesnelheid!
daarom, als de meeste mutaties neutraal zijn (zoals voorgesteld in de neutrale theorie) en als de mutatiesnelheden constant zijn in de tijd, moeten substituties ook constant in de tijd voorkomen. We moeten dan klok-achtige regelmatige snelheden van substituties observeren op moleculair niveau. Kimura (1969) beschouwde de observatie van relatief constante moleculaire klok in eiwitsequenties als een sterke ondersteuning voor de neutrale theorie van moleculaire evolutie.
testen hoe de neutrale moleculaire klok loopt
volgens de neutrale theorie is de vraag of substitutiesnelheden constant zijn in de tijd gelijk aan of neutrale mutatiesnelheden constant zijn in de tijd. Om deze reden, richtten vele latere studies zich op het analyseren van gegevens van neutrale plaatsen om te bepalen of neutrale mutatietarieven inderdaad constant in de tijd zijn. We zullen kort bespreken hoe deze studies worden uitgevoerd, voordat we theoretische debatten over moleculaire klokken bespreken. De Analyses van eiwitmoleculaire klokken werden ook voortgezet, maar de debatten rond variatie in eiwitmoleculaire klokken verschillen sterk van die over neutrale moleculaire klokken, en zullen niet in dit artikel worden opgenomen. Geïnteresseerde lezers zouden Gillespie (1991), Kumar (2005), Kim and Yi (2008) en Bedford et al. (2008). de meeste empirische analyses van neutrale moleculaire klokken baseren zich op de stelling dat neutrale mutatiesnelheden kunnen worden afgeleid uit neutrale substitutiesnelheden (Kimura 1968, 1969). In de praktijk definieert elke studie een bepaald type van plaatsen in het genoom als neutrale plaatsen, en vergelijkt substitutiepercentages van die plaatsen tussen geslachten.
welke plaatsen in het genoom echt neutraal zijn kan niet volledig worden bepaald, maar wetenschappers kwamen met een aantal nuttige proxies. Vóór het tijdperk van genoom het rangschikken, waren de meeste beschikbare opeenvolgingsgegevens die van eiwit-codeert opeenvolgingen van DNA. Studies verdelen vaak eiwitcoderende DNA-sequenties in twee typen locaties (Wu & Li 1985). Het eerste type sites omvat sites waarvoor elke verandering zou leiden tot aminozuursubstituties, of ‘niet-synonieme sites’. Het tweede type sites, ‘synonieme sites’, omvat sites die ‘gedegenereerde’ posities coderen in de codontabel, waar een verandering niet leidt tot een aminozuursubstitutie. Bijvoorbeeld, TCT en TCC coderen beiden serine. Als de derde positie van deze codon verandert, zullen ze nog steeds dezelfde aminozuren produceren. Dergelijke substituties zouden minder zichtbaar zijn voor natuurlijke selectie. Daarom moeten moleculaire klokken op synonieme locaties dichter bij de neutrale moleculaire klok staan dan niet-synonieme klokken.
aangezien sequentietechnieken geavanceerd waren, gebruikten sommige studies introns als proxies voor neutrale locaties, omdat ze niet zijn opgenomen in de volwassen mRNA ‘ s, en dus eerder neutraal zijn (Yi et al. 2002). Sequenties van inactieve transposable elementen die werden ingevoegd lang voordat species divergentie werden ook vaak gebruikt (deze worden vaak aangeduid als ‘voorouderlijke herhalingen’, bijvoorbeeld, Thomas et al., 2003). Tot slot gebruikten sommige studies niet-coderende DNA-sequenties (alle sequenties na het verwijderen van eiwitcoderende DNA-sequenties) uit gehele genoomuitlijningen om neutrale moleculaire klokken te testen (Elango et al. 2006, Huttley et al. 2007).
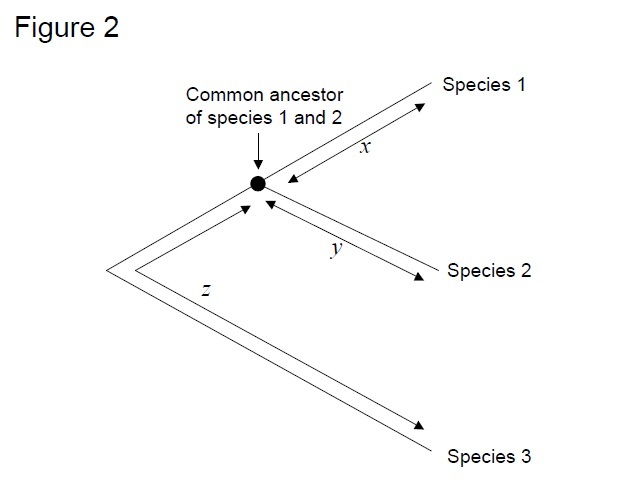
de meest gebruikte test is de zogenaamde “relative rate test”(Sarich & Wilson 1973). Aanvankelijk werden substitutiepercentages per tijdseenheid geschat door het totale aantal verschillen (substituties) tussen eiwitten van verschillende soorten te delen door de divergentietijd, geschat op basis van fossiele records (figuur 1). Fossiele records zijn echter niet beschikbaar voor veel vergelijkingen en worden geassocieerd met grote foutmarges. De relatieve snelheid test overwint de noodzaak voor fossiele records (Figuur 2). Zolang er een uitgroepsequentie bestaat naar de twee interestlijnen, kunnen we bepalen of de twee takken dezelfde of verschillende moleculaire klokken volgen, zonder de kennis van de absolute tijd van divergentie (Figuur 2).
Wat zijn de determinanten van neutrale moleculaire Klokken?
bijna alle controverses die centraal staan in debatten over neutrale moleculaire klokken komen voort uit de vraag wat de belangrijkste bronnen van mutaties zijn. Deze vraag is direct relevant voor het begrijpen van patronen van mutatie, die de ultieme bron van evolutionaire verandering en genetische ziekte zijn. Voorts is het begrijpen hoe de veranderingstarieven tussen geslachten en binnen genomen variëren een fundamentele vraag in vergelijkende genomica, die sequentievergelijkingen beoogt te gebruiken om genomic gebieden te identificeren die functioneel belangrijk zijn.
dus wat bepaalt de neutrale mutatiepercentages? Een van de belangrijkste bijdragen aan neutrale moleculaire klokken is lineage-specifieke variatie in generatietijden. Vanaf het begin werd het idee van een constante neutrale moleculaire klok gezien als zijnde in strijd met de moleculaire mechanismen van germline mutatie. Men heeft lang overwogen dat de meeste veranderingen uit fouten in DNA-replicatie in kiemlijnen voortvloeien (Haldane 1947, Muller 1954). Aangezien de veranderingen voorkomen wanneer germline DNA voor de volgende generatie wordt herhaald, zouden zij in verhouding tot het aantal generaties, eerder dan de absolute hoeveelheid tijd moeten accumuleren. Daarom, als we de aantallen substituties vergelijken die zich sinds hun divergentie in twee geslachten hebben opgehoopt, zou de afstamming met langere generatietijd, die minder DNA-replicatie gebeurtenissen heeft ondergaan, minder substituties bevatten in vergelijking met de afstamming met de kortere generatietijd. Daarom moet de moleculaire klok langzamer lopen bij soorten met langere generatietijden. Dit idee wordt aangeduid als het’generation-time effect’.
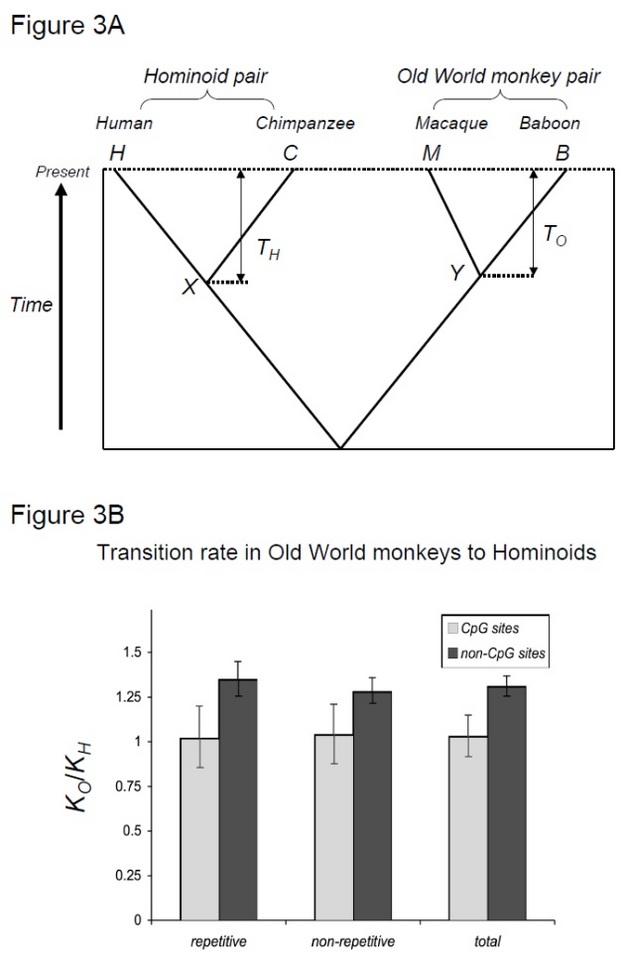
in feite werd het generatietijd-effect voor het eerst waargenomen in studies van primaten, zelfs vóór het debat over moleculaire klok. Morris Goodman, die op dat moment immunologische methoden gebruikte om soortverwantschap te onderzoeken, merkte op dat de snelheid waarmee sommige eiwitten uiteenlopen bij apen, in het bijzonder bij mensen, lijkt te zijn afgenomen in vergelijking met apen uit de Oude Wereld (Goodman 1961, 1962, 1963). Dit effect wordt aangeduid als’hominoide rate slowdown’. Aangezien hominoôden langere generatietijden hebben dan apen van de oude wereld, kan deze observatie worden verklaard door het generatietijd-effect.
Wu & Li (1985) testten als eerste de generatietijd-effecthypothese met behulp van DNA-sequentiegegevens. Ze gebruikten gegevens van 11 genen van primaten en knaagdieren. Aangezien primaten een veel langere generatietijd hebben dan knaagdieren, zou de moleculaire klok bij knaagdieren sneller moeten zijn dan bij primaten. Zij stelden namelijk vast dat knaagdieren voor synonieme locaties ongeveer twee keer zo snel evolueren als primaten (Wu & Li 1985). Voor niet-synonieme sites werd een dergelijk effect echter niet gevonden. Met andere woorden, de neutrale moleculaire klok, maar niet de aminozuur moleculaire klok, tikt sneller in de knaagdier Lijn in vergelijking met de primaten lijn, die goed past bij het idee van een generatie-tijd effect.
latere studies leverden verdere ondersteuning op voor de vertraging van de hominoôdesnelheid (Li & Tanimura 1987, Bailey et al. 1991) en het snelheidsverschil tussen de knaagdieren en de primaten (Gu & li 1992, Huttley et al. 2007). Bovendien werd een snelheidsverschil waargenomen in nog kleinere fylogenetische schalen, vooral bij primaten. : bijvoorbeeld, de menselijke moleculaire klok loopt langzamer dan de chimpansee moleculaire klok (Elango et al. 2006); de percentages bij apen van de nieuwe wereld zijn hoger dan die bij mensachtigen en apen van de Oude Wereld (Steiper & Young 2006). De verschillende snelheden van moleculaire klokken waargenomen in deze studies zijn kwalitatief consistent met het generatietijd-effect.
de werkelijke verschillen tussen geslachten zijn echter niet kwantitatief consistent met het verschil in generatietijd. Kumar & Subramanian (2002) toonde bijvoorbeeld aan dat hoewel het verschil in generatietijd tussen primaten en knaagdieren veel groter is dan dat tussen mensen en apen uit de Oude Wereld, de waargenomen verschillen in moleculaire klokken in deze twee vergelijkingen vergelijkbaar zijn. Het is de moeite waard op te merken dat Kumar & Subramanian (2002) specifieke statistische filters gebruikte om gegevens te verwijderen die ‘heterogene’ substitutiepatronen vertoonden, die mogelijk een vertekening naar langzaam evoluerende sequenties hebben veroorzaakt (Yi et al. 2002). Niettemin lijkt het verschil in moleculaire klokken van primaten en knaagdieren veel kleiner dan oorspronkelijk voorgesteld door Wu & Li (1985). Bijvoorbeeld, Huttley et al. (2007) analyseerde gehele genoomuitlijningen van verschillende soorten, waaronder de opossum, en toonde aan dat het tariefverschil tussen eutherische lijnen en de opossum lijn (~30%) veel groter is dan het tariefverschil tussen mens en muis lijnen (~14%). Deze voorbeelden tonen aan dat de mate van verschillen in moleculaire klokken aanzienlijk varieert tussen verschillende studies als gevolg van verschillen in datasets en statistische methoden. Zij tonen ook aan dat tariefverschillen tussen geslachten niet volledig kunnen worden verklaard door het verschil in generatietijd alleen. Er zijn duidelijk andere bijdragen aan neutrale moleculaire klokken.
inderdaad, kenmerken van de levensgeschiedenis anders dan generatietijden lijken te variëren met moleculaire klokken. Martin & Palumbi (1993) toonde aan dat DNA moleculaire klokken sneller lopen bij soorten met een kleine lichaamsgrootte. Deze observatie leidde tot de hypothese dat metabolische snelheden belangrijke determinanten van moleculaire klokken zijn. Een hoog metabolisme veroorzaakt grote aantallen mutagene zuurstofradicalen, die mutatiepercentages zouden verhogen (Rand 1994). Omdat de metabolische tarieven en lichaamsgrootte over het algemeen mede variëren met de generatietijden, is het moeilijk geweest om te onderscheiden welke van deze de belangrijkste determinant van moleculaire kloksnelheden vormt. Tsantes & Steiper (2009) hebben op basis van gegevens van primaten voorgesteld dat leeftijd bij de eerste voortplanting, in plaats van lichaamsgrootte, de belangrijkste determinant is van moleculaire klokken. Aangezien de leeftijd bij eerste reproductie het effect van de generatietijd weerspiegelt, ondersteunt deze studie het idee dat het effect van de generatietijd de belangrijkste determinant van moleculaire klok is. Deze studie is echter nog steeds gebaseerd op een beperkt aantal geslachten (er werden vier paren soorten gebruikt). Aldus, onderscheidend tussen lichaamsgrootte, generatie-tijd effect, en metabolische tarieven blijft nog steeds als een belangrijke kwestie in het generaliseren en begrijpen van neutrale moleculaire klokken.
bovendien wordt het belang van factoren die niet met de generatietijd variëren, zoals DNA-methylering, steeds meer gewaardeerd. Methylation van DNA is een chemische die wijziging van genomic DNA in diverse taxa wordt gevonden. In dierlijke genomen, DNA methylering gebeurt bijna uitsluitend bij cytosines gevolgd door guanines (zogenaamde ‘CpG’ s). Geméthyleerde cytosines hebben op hun beurt de neiging om snel te muteren in thymines als gevolg van chemische instabiliteit (Bird 1980). In het menselijk genoom komen mutaties veroorzaakt door DNA methylering meer dan een orde van grootte vaker voor dan andere mutaties (Nachman & Crowell 2000, Elango et al. 2008). Omdat de veranderingen die door methylation van DNA worden veroorzaakt grotendeels onafhankelijk van de replicatie van DNA voorkomen, kunnen dergelijke veranderingen verschillende moleculaire klokken volgen dan anderen. Specifiek, in plaats van generatie-tijd afhankelijkheid, mutaties veroorzaakt door DNA methylering kunnen volgen een tijd-afhankelijke moleculaire klok, die vergelijkbaar is met wat aanvankelijk werd voorgesteld door Zuckerkandl & Pauling (Kim et al. 2006)!
om deze hypothese te testen, Kim et al. (2006) vergeleken divergentie tussen mens-chimpansee en divergentie tussen makaak-baviaan, twee soorten paren die dezelfde divergentie tijden delen maar met verschillende generatietijden (Steiper et al. 2004). Het mens-chimpanseepaar (het hominoidepaar) heeft veel langere generaties in vergelijking met het makaak-baviaan-paar (het Oude-Wereld-apenpaar). Deze studie toonde aan dat Voor niet-CpG sites, de Oude Wereld apenpaar geaccumuleerd ongeveer 30% meer substituties, die kan worden verklaard door de bovengenoemde hominoïde snelheid vertraging effect. In tegenstelling, moleculaire klokken op CpG-sites vertoonden vergelijkbare aantallen substituties in hominoïde en oude wereld aap paren (Figuur 3). Aldus, bestaan tijd-afhankelijke en generatie tijd-afhankelijke moleculaire klokken naast elkaar binnen dezelfde genomen. De veronderstelling dat een enkele moleculaire klok kan bestaan voor een bepaalde lijn is niet langer geldig, omdat de overheersende mutatiekrachten variëren tussen genomische gebieden.
conclusies
het concept van een constante moleculaire klok werd aanvankelijk voorgesteld op basis van een beperkte hoeveelheid gegevens over de eiwitsequentie. Hoewel latere studies aantoonden dat een dergelijke observatie geen algemeen patroon in aminozuren is, heeft het een significante invloed gehad op het gebied van moleculaire evolutie, in het bijzonder op de ontwikkeling van de neutrale theorie van moleculaire evolutie voor DNA-sequentiegegevens. Volgens de neutrale theorie richtten de studies zich op het ophelderen van patronen van variatie in neutrale mutatiesnelheden. Gedurende de laatste decennia hebben we vastgesteld dat moleculaire klokken op verschillende snelheden lopen tussen de geslachten. Bovendien kan de mate van variatie variëren afhankelijk van de verschillende soorten gegevens en specifieke statistische methoden die worden gebruikt. Het generatietijd-effect blijft op een kwalitatief niveau gehandhaafd, maar is onvoldoende om de kwantitatieve variatie van neutrale mutatiesnelheden tussen geslachten te verklaren. De eigenschappen van de levensgeschiedenis en de niet-replicatie afhankelijke veranderingen, zoals die door methylation van DNA worden veroorzaakt, zijn ook belangrijke bijdragers aan genomic moleculaire klokken. Inderdaad, worden de verschillende types van moleculaire klokken waargenomen zelfs binnen een genoom, omdat de overheersende mutationele input tussen verschillende genomic gebieden varieert. Dus, in plaats van het aannemen van een enkele neutrale moleculaire klok voor elk genoom, toekomstige studies moeten gericht zijn op het onthullen van de variatie van genomische neutrale moleculaire klokken, om te leren over genomische mutationele Landschappen. Dergelijke informatie is niet alleen nuttig voor het begrijpen van de grondstof die moleculaire evolutie en genetische ziekte regeren, maar vormt ook een kritische component die vergelijkende en functionele genomic analyses beà nvloedt om functionele genomic gebieden te identificeren.