Inleiding
Internationaal instandhoudingsbeleid erkent biodiversiteit op drie niveaus: ecosysteem, soorten en genetisch, en dat Beheer ernaar moet streven alle drie te behouden (Verdrag inzake biologische diversiteit, 2007). Dit komt duidelijk tot uiting in het Verdrag inzake biologische diversiteit, Aichi Biodiversity Targets, dat in 2010 is overeengekomen (https://www.cbd.int/sp/targets/), waarin in doelstellingen en streefdoelen niet alleen naar ecosystemen en soorten, maar ook naar genetische diversiteit wordt verwezen. Toch zijn de huidige benaderingen van het behoud van biodiversiteit grotendeels gebaseerd op geografische gebieden, ecosystemen, ecologische gemeenschappen en soorten, met minder aandacht voor genetische diversiteit en het soortenpopulatiecontinuüm. De meerderheid van de natuurbeschermers richt zich op soorten als maatstaf voor biodiversiteit, met enige aandacht voor intraspecifieke eenheden binnen intensief beheerde bedreigde soorten. In deze context is er een natuurlijk verlangen om eenvoudige categorisaties te gebruiken—benoemde soorten, en andere benoemde taxa (dat wil zeggen, ondersoorten), en soms instandhoudingseenheden binnen soorten. Soorten in het bijzonder bieden een gemeenschappelijke maat voor biodiversiteit die veel wetenschappelijke en beheersinspanningen ondersteunt (Mace, 2004; Wilson, 2017). Als fundamentele eenheid van instandhouding worden soorten vaak gebruikt om de waarde van de biodiversiteit te kwantificeren door middel van lijsten van soorten binnen beschermde gebieden, identificatie van bedreigde soorten binnen rechtsgebieden en als basis voor bioveiligheidsprocedures. Daarnaast zijn soorten typisch de entiteiten waarmee we instandhoudingsproblemen communiceren aan het publiek.
deze impuls om eenvoudig discrete eenheden in de natuur te identificeren, druist in tegen verschillende, verwante realiteiten. Ten eerste is speciatie niet onmiddellijk. Eerder is het typisch een langdurig proces (Dynesius and Jansson, 2014; Roux et al., 2016), voortschrijdend van geïsoleerde populaties, misschien onderworpen aan verschillende selectiedruk, tot eenduidige soorten over lange tijdschalen, met frequente omkeringen veroorzaakt door omgevingsverandering of eenvoudige genetische samenvoeging als bereiken opnieuw aansluiten (figuur 1; Seehausen et al., 2008; Rosenblum et al., 2012). Ten tweede zijn de meeste erkende soorten niet genetisch uniform. Divergentie (drift en langdurige isolatie) en reticulatie (geneflow, hybridisatie, introgressie en recombinatie) vormen effectief geografische en genomische variatie binnen soorten (Cutter and Payseur, 2013; Edwards et al., 2016) en sommige soorten kunnen worden gestructureerd in lange geïsoleerde populaties, waarvan sommige erkenning als volledige maar misschien morfologisch cryptische soorten zouden kunnen rechtvaardigen (Bickford et al., 2007; Jorger en Schrödl, 2013). Ten derde, zo lang erkend, benoemde soorten zijn niet gelijkwaardig in termen van hoeveel evolutionaire erfenis ze vertegenwoordigen—simpel gezegd, een vogelbekdier (als vertegenwoordiger van een oud uiteenlopend geslacht) kan worden beschouwd als meer waard dan een onlangs uiteengelopen knaagdier (Isaac et al., 2007). Ten vierde worden soortaanduidingen niet in steen gegoten, maar zijn het hypothesen die moeten worden getest als er nieuw bewijs beschikbaar komt (Hey et al., 2003). Elk van deze kwesties creëert complexiteit en dubbelzinnigheid bij het afbakenen van soorten in het algemeen, en het toekennen van aandacht voor behoud aan hen of hun samenstellende populaties. Daarnaast is de afbakening van soorten ook onderworpen aan verschillende soorten concepten en manieren van diagnosticeren, die grote consternatie heeft veroorzaakt onder conservatie biologen (Mace, 2004; Frankham et al., 2012; Garnett and Christidis, 2017).
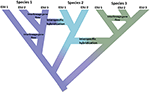
figuur 1. Speciatie als continuüm van geïsoleerde populaties tot eenduidige soorten, zoals blijkt uit de kleurverschuiving waarbij de kleuren verschillende soorten vertegenwoordigen. Sommige geslachten die ontstaan als gevolg van populatieisolatie ontwikkelen zich tot echte soorten met de interne knooppunten van de boom die initiatie van het speciatieproces vertegenwoordigen. Andere geslachten ontwikkelen zich niet tot echte soorten, maar verschillen in de tijd, wat erop wijst dat soorten niet genetisch uniform zijn. Deze geslachten worden weergegeven als evolutionair significante eenheden (ESUs). Inter-lineage gen flow is geïndiceerd voor Species 1 en Species 3. In dit geval kan evolutionaire onafhankelijkheid van voorbijgaande aard zijn omdat twee geslachten die tot dezelfde soort behoren niet reproductief geïsoleerd zijn. Terwijl zij als verschillende geslachten (soorten) blijven, kan interspecifieke hybridisatie met beperkte genuitwisseling (verschuiving in kleur) tussen species voorkomen.
implementatie van effectieve instandhoudingsstrategieën die gericht zijn op de bescherming van zowel soorten als genetische diversiteit zou baat hebben bij zowel een consistentere visie op hoe we soorten en andere eenheden in het populatie-soortencontinuüm afbakenen, als hoe deze eenheden wereldwijd in verschillende wetgeving worden geïnterpreteerd. Nieuwe mogelijkheden van genomica bieden zowel kansen als uitdagingen om deze doelen te bereiken. In het volgende overwegen we: (i) hoe genoom-schaal analyses kan worden verzoend met een conservatieve aanpak voor de afbakening en naamgeving van de soorten, terwijl ook het identificeren van de belangrijkste onderdelen van de genetische diversiteit binnen soorten, en (ii) hoe behoud bestaande wetgeving op verschillende continenten, die bepaalt “soorten” te omvatten een breed scala van infra-specifieke categorieën kunnen worden gebruikt, gezien de hoge resolutie aangeboden door genoom schermen, en (iii) hoe de inzichten in processen van soortvorming en de structurering van de variatie van genomics in lijn zijn met de wetgeving en vervolgens in het management. Vervolgens stellen we een aantal manieren voor om de instandhoudingseenheden op één lijn te brengen met de wetgevingsbenaderingen om het volledige spectrum van diversiteit van populaties tot soorten te beschermen.
Genomica, soortvorming en taxonomie
net als vele gebieden van de biologische wetenschap, wordt systematiek, de praktijk van het benoemen van taxa, en het begrijpen van hun relaties en hoe ze vormen, aanzienlijk bevorderd door genomica en statistische modellen. Of door hele genoom sequencing (bijv., Nater et al., 2017) of genoombemonstering (SNPs, target capture-sequencing; Jones and Good, 2016; Leache and Oaks, 2017), hebben we nu een ongekende resolutie van patronen van genetische diversiteit (Funk et al., 2012). In combinatie met steeds krachtiger statistische modellen voor het afleiden van geschiedenissen van genomische en species divergentie, bieden deze gegevens nieuwe inzichten in de evolutionaire processen die soorten en genetische variatie genereren.
toepassing van deze nieuwe tools genereert twee tegengestelde inzichten. Aan de ene kant, veel studies onthullen zeer uiteenlopende genetische populaties binnen genoemde soorten, zogenaamde cryptische soorten (Bickford et al., 2007; Jorger en Schrödl, 2013; Hitch et al., 2017). Dit is niets nieuws, want het bouwt voort op een lange geschiedenis van ruimtelijke populatiegenetica en moleculaire fylogeografie die al drie decennia een productieve onderneming is (Avise et al., 1987). Wat genomica tot dit streven brengt is zowel een verhoogde resolutie van de populatiestructuur Als gegevens die voldoende zijn om statistisch populatiegeschiedenissen af te leiden en alternatieve divergentiemodellen te testen (Degnan and Rosenberg, 2009; Sukumaran and Knowles, 2017). Aan de andere kant tonen analyses op genoomschaal ook aan dat (zoals al lang erkend voor planten) genetische uitwisseling (introgressie) tussen diersoorten vaker voorkomt dan eerder gedacht, zowel tijdens als na speciatie, en zelfs nieuwe adaptieve straling kan aandrijven (Mallet, 2007; Rieseberg and Willis, 2007; Arnold and Kunte, 2017) figuur 1. Samen benadrukken deze inzichten verder dat speciatie langdurig is (Roux et al., 2016) en onthullen de tegengestelde krachten die werken door het fylogeography-phylogenetics continuüm (Edwards et al., 2016).
hoe sluit dit alles aan bij taxonomie en instandhoudingsdoelstellingen? Er is sterke en gerechtvaardigde bezorgdheid dat de verhoogde resolutie geboden door genoomschermen zou kunnen leiden tot welig tierende over-splitsing van soorten (Isaac et al., 2004), mogelijk het beperken van management flexibiliteit en het overdragen van kleine genetisch uiteenlopende populaties inteelt en uiteindelijke uitsterven (Frankham et al., 2012). Omgekeerd, Gippoliti et al. (2018) argumenteren dat taxonomische inertie, resulterend in eerdere niet-erkenning van veel soorten Afrikaanse hoefdieren, had geresulteerd in een achterblijvende depressie na vermenging en het niet-beschermen van geografisch beperkte taxa. Hun mening (en zie ook Morrison et al., 2009) is dat er geen bewijs is dat taxonomische inflatie negatieve effecten heeft op het behoud van hoefdieren (Gippoliti et al., 2018). Een daarmee verband houdende zorg is dat onstabiele taxonomie zowel de praktijk van conservering als de publieke percepties vertraagt (Garnett and Christidis, 2017). Deze bezorgdheid wordt deels veroorzaakt door het langdurige debat rond soortconcepten en diagnostische methoden in de systematische biologie, in het bijzonder de toepassing van het fylogenetische Soortenconcept dat stelt dat soorten minimaal diagnostische eenheden zijn (Cracraft, 1983). Echter, de meerderheid van de taxonomisten zijn inherent conservatief, alleen het benoemen van soorten als er overeenstemming is over meerdere lijnen van bewijs (Dayrat, 2005; Padial et al., 2010; Schlick-Steiner et al., 2010: Yeates et al., 2011). Ons standpunt is dat soorten robuuste onafhankelijk evoluerende geslachten moeten vertegenwoordigen die grotendeels intact blijven wanneer ze in contact komen met naaste verwanten—in overeenstemming met de bedoeling van het evolutionaire Soortenconcept (Wiley, 1978) en het gegeneraliseerde Soortenconcept (de Queiroz, 1998, 2007). In de praktijk vereist dit direct bewijs van reproductieve isolatie of redelijke surrogaten van hetzelfde, terwijl beperkte genetische uitwisseling mogelijk is (Singhal et al., 2018).
genomische gegevens bieden meer vermogen dan voorheen om differentiatie en divergentie binnen en tussen soorten te identificeren, waardoor meer aandacht nodig is voor de manier waarop dit de afbakening van taxonomische eenheden beïnvloedt. Waar populaties zowel sterk genetisch als fenotypisch uiteenlopen ten opzichte van reeds genoemde soorten, is er een sterke reden om deze als afzonderlijke soorten te benoemen. Als populaties fenotypisch sterk uiteenlopen met weinig genomische scheiding, dan is er een beoordelingsvraag of deze fenotypische verschillen (i) erfelijk zijn, en (ii) waarschijnlijk substantiële reproductieve isolatie veroorzaken. Het aftasten van het genoom kan dergelijke situaties oplossen door voor beperkte genstroom te testen. De meer uitdagende kwestie is waar morfologisch vergelijkbare, maar genetisch uiteenlopende populaties worden gedetecteerd. Dergelijke populaties kunnen gemakkelijk worden gediagnosticeerd als aparte evolutionaire lijnen door statistische afbakeningsmethoden toe te passen op multilocus data (Carstens et al., 2013; Rannala, 2015), maar zijn het vluchtige populaties of duurzame soorten (Sukumaran and Knowles, 2017)? Het is zeker mogelijk, binnen de regels van de nomenclatuur, om soorten te herkennen uit DNA-sequenties alleen, het verstrekken van sequentie-gebaseerde diagnostiek (Tautz et al., 2003; Cook et al., 2010). Bijvoorbeeld, Murphy et al. (2015) delimit en beschrijven zes Australische woestijn voorjaar amfipod soorten op basis van DNA sequentie data ondanks geen duidelijke morfologische verschillen. Echter, dit is een algemeen gebied voor verbetering waar systematisten nodig hebben om de term “cryptische soorten” toe te passen met meer consistentie en rigor (hitched et al., 2017).
in situaties waarin DNA-sequentiegegevens een mogelijk sleutelbepalend kenmerk vormen voor de herkenning van een soort, stellen we voor dat het vooral belangrijk is om andere aanwijzingen toe te passen om oversplitting te voorkomen (Oliver et al., 2015). Een methode is om te testen op substantiële reproductieve isolatie waar dergelijke populaties natuurlijke contactzones vormen. Bijvoorbeeld, in een vergelijkende genomische analyse van contactzones onder geslacht-paren van Carlia hagedissen die fenotypisch niet te onderscheiden zijn, werden indices van genetische isolatie sterk gecorreleerd met divergentietijd (Singhal and Moritz, 2013; Singhal and Bi, 2017) (Figuur 2). De meer divergente van deze afstamming-paren zijn nu formeel erkend als afzonderlijke soorten, terwijl meer nauw verwante (maar statistisch diagnostische) lijnen met bewijs van genomisch uitgebreide genetische uitwisseling tussen contactzones waren niet (Singhal et al., 2018). Gezien de sterke empirische relatie tussen divergentie en reproductieve isolatie in deze taxa, deze auteurs verder gescheiden twee allopatric en cryptische isolaten (van een nauw verwante soort) die nog hogere genomische divergentie dan die hierboven afgebakend. Uitbreiding van deze reproductieve isolatie informed metric naar andere congeneren soorten met diep uiteenlopende maar morfologisch vergelijkbare fylogeografische lijnen (Potter et al., 2016, 2018; Afonso Silva et al., 2017a) suggereert dat er nog meer soorten moeten worden beschreven, vooral op eilanden (Figuur 2, Carlai amax en C. rufilatus), en inderdaad, sommige van dergelijke soorten zijn beschreven na complementaire fenotypische analyse (Afonso Silva et al., 2017b). Andere afstammelingen met recentere verschillen, maar nog steeds statistisch gescheiden als afzonderlijke afstammelingen met behulp van een grote reeks nucleaire genen, kunnen worden weergegeven als evolutionair significante eenheden (ESUs) binnen soorten.
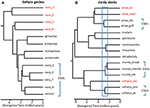
Figuur 2. Fylogenomic resolutie van soortgrenzen in hagedissen uit de Australische Moessoonse tropen (AMT). In elk geval worden relaties en relatieve divergentietijden geschat op basis van 100 exons met behulp van de multispecies coalescent geïmplementeerd in StarBEAST2, maar de tijdschalen tussen de twee voorbeelden zijn niet noodzakelijk hetzelfde. A) resultaten voor de Nana-groep Gehyra-gekko ‘ s (Moritz et al., 2018). De fylogenetische diversiteit van geslachten binnen G. nana, zoals eerder erkend, omvatte vier andere reeds beschreven en morfologisch onderscheidende soorten. Latere analyses van range overlappingen en morfologie hebben geleid tot herkenning van elk van de meest fylogenetisch divergente geslachten (rode taxon labels) als afzonderlijke soorten (Doughty et al., 2018). De (morfologisch onafscheidelijke) afstammelingen die in G. nana achterblijven, kunnen als ESUs worden beschouwd. (B) doorlopend oplossen van de soortgrenzen in Carlia skinks van het AMT. Sommige geslachten zijn al erkend als soort op basis van een combinatie van genetische soortafbakening en post-hoc identificatie van diagnostische morfologische verschillen (C. insularis en C. isostriacantha; Afonso Silva et al., 2017b). Voor andere complexen (Potter et al., 2016, 2018), zijn taxonomische herzieningen aan de gang. De blauwe rechthoek geeft het bereik van divergentie tijden waar aanzienlijke reproductieve isolatie, gecorreleerd met divergentie, is waargenomen in contactzones tussen morfologisch cryptische geslachten van verwante soorten (Singhal and Bi, 2017). Conservatief, en volgende (Singhal et al., 2018), erkennen we als kandidaat-soort drie genetische afstamming met verschillen groter dan deze empirische drempel. Dit zou genetisch verschillende ESUs binnen elk van Carlia amax, Carlia munda, en Carlia rufilatus (blauwe haakjes). De schaalbalken vertegenwoordigen schattingen van de divergentietijd in miljoenen jaren.
in extreme gevallen blijkt uit genomische methoden dat één taxonomische soort niet alleen genetisch heterogeen is, maar in fylogenetische zin ook andere benoemde soorten bevat. Een dergelijke” paraphyly ” kan ontstaan hoewel recente speciatie van een enkele, genetisch onderverdeeld soort (Patton and Smith, 1994), maar het genereert ook de hypothese dat het genetisch heterogene taxon meerdere soorten kan vertegenwoordigen. Nogmaals, er is meer bewijs nodig. Fylogenomic analyse van een complex van Gehyra gecko hagedissen uit Noord-Australië scheidde negen evolutionaire lijnen af, met vier reeds benoemde soorten fylogenetisch ingebed onder hen (Moritz et al., 2018; Figuur 2). Later bewijs van geografische overlapping (met behoud van fylogenetische scheiding) en subtiele morfologische verschillen ondersteunde de naamgeving van de vier meest fylogenetisch divergente geslachten (Doughty et al., 2018). Omgekeerd hadden de nauw verwante (maar statistisch scheidbare) geslachten aangrenzende in plaats van overlappende distributies, maar waren morfologisch niet diagnosticeerbaar, en werden bijgevolg behouden als een enkele soort (G. Nana sensu stricto), zij het met meerdere ESUs, in afwachting van verder onderzoek (Figuur 2). Het volgende genoomaftasten van steekproeven over de grenzen van deze ESUs onthulde dat minstens wat wezenlijke genetische uitwisseling hebben, het valideren van de originele, conservatieve benadering die voor het erkennen van species wordt genomen.
in andere gevallen kunnen plantengenomische studies, zelfs door eenvoudige bevestiging van SNP ‘ s, sterke en onverwachte verschillen tussen populatiemonsters aan het licht brengen, wat wijst op de aanwezigheid van cryptische taxa die vervolgens kunnen worden geanalyseerd op diagnostische fenotypische karakters. Bijvoorbeeld, SNP-gebaseerd bewijsmateriaal voor genomic divergentie tussen populaties en ecotypic verschillen in Eucalyptus salubris onthulde twee verschillende moleculaire lijnen die hun onderscheidend vermogen in een gebied van geografische overlap handhaafden, met ecotypic aanpassing beschouwd als een belangrijke factor in het minimaliseren van genstroom tussen de lijnen. (Steane et al., 2015; Figuur 3). In een ander geval, in Australisch Pelargonium, heeft genomische analyse, gecombineerd met morfologische en reproductieve studies, geleid tot significante veranderingen in taxonomie, zowel het splitsen en klonteren van taxa, vooral waar eerder gebruikte morfologische eigenschappen labiel en disharmonisch met fylogenie blijken te zijn (Nicotra et al., 2016).
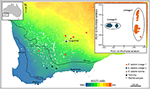
Figuur 3. Genomische analyse met behulp van SNP-gegevens identificeerde cryptische lijnen in de wijdverspreide Eucalyptus salubris. De plaats van steekproevenpopulaties van elke lijn wordt getoond over een aridity index gradiënt in het zuidwesten van West-Australië. PCoA gebaseerd op binaire genetische afstanden afgeleid van 16.122 dartseq tellers, toont twee verschillende geslachten. Aangepast van Steane et al. (2015).
de bovenstaande gevallen illustreren de progressieve hypothese-testing benadering die is gebruikt in integratieve taxonomie (Padial et al., 2010; Yeates et al., 2011). Verder, gebruiken deze gevallen één of andere vorm van genoomreductie rangschikkend om hoogte-kwaliteitsgegevens van 1000 ‘ s van onafhankelijke genen te verkrijgen, die veel groter vertrouwen in de resultaten geven dan het geval gebruikend enkel plastid loci of een paar nucleaire genen is. Een voorbeeld van hoe het toepassen van integratieve taxonomie de percepties van diversiteit drastisch kan veranderen, komt uit een gecombineerd genetisch, akoestisch en morfologisch onderzoek van kikkers uit Madagaskar dat de bekende soortendiversiteit bijna verdubbelde (Vieites et al., 2009). Deze auteurs stelden een reeks termen voor die de staat van taxonomische resolutie en congruentie weerspiegelen: Bevestigde kandidaat-soorten, voor genetisch en fenotypisch divergente, maar nog niet met de naam taxa; onbevestigde kandidaat-soorten voor genetisch divergente taxa waarvan de morfologie en akoestiek nog niet waren onderzocht; en diep Conspecifieke afstamming voor entiteiten waarvan bekend is dat zij genetisch divergente maar fenotypisch cryptische entiteiten zijn. Genomische analyses moeten nog worden toegepast op deze taxa, maar zouden zeker beslissingen over conspecific lineages vs. kandidaat-soorten helpen.
genomische benaderingen, zelfs wanneer conservatief toegepast, zullen ongetwijfeld leiden tot verdere splitsing van morfologisch conservatieve taxa. Maar, omgekeerd, zijn er duidelijke voorbeelden waar dezelfde benaderingen onthullen over-split of onjuist gediagnosticeerde soorten (Nicotra et al., 2016). Natuurlijk zijn er al veel moleculaire studies op plant-en diersoorten complexen gebaseerd op kleinere datasets (bijv., microsatelliet, mtDNA, en cpDNA) die zijn geïdentificeerd waar soorten zijn verkeerd gediagnosticeerd, of waar meerdere soorten zijn omschreven taxonomisch, maar worden nu beschouwd als een enkele soort. Bijvoorbeeld, Rabosky et al. (2014) gebruikte mitochondriale en nucleaire genfylogenieën om het scincid ctenotus inornatus complex te herzien, resulterend in een netto vermindering van soorten en herschikking van soortgrenzen; de morfologische eigenschappen die eerder werden toegepast, terwijl geschikt voor veldbiologen, waren zeer labiel en grotendeels niet aan soortgrenzen zoals genetisch gediagnosticeerd overeenkomen. In planten zijn enkele van de meest bekende voorbeelden te vinden in de orchideeën, zoals de Europese orchideeëngeslacht Ophrys en de Australische orchideeëngeslacht Caladenia, waar moleculaire studies wijzen aanzienlijk minder entiteiten die zouden worden erkend als soorten, dan die op basis van morfologische eigenschappen en pseudocopulatory bestuiving syndromen (zie Devey et al., 2008; Swarts et al., 2014).
gegevens op de Genoomschaal, indien conservatief en idealiter geïnterpreteerd met parallelle analyses van fenotypen, zullen blijven leiden tot taxonomische veranderingen, vooral in morfologisch conservatieve taxa (Fišer et al., 2018). Maar, in tegenstelling tot de zorgen geuit door sommigen (Garnett en Christidis, 2017), We zijn van mening dat dit het juiste nastreven van evolutionaire systematiek, zelfs als de resultaten zijn tijdelijk onhandig.
Genomica en intraspecifieke Instandhoudingseenheden
hoewel minder controversieel dan taxonomische afbakening en het debat op soortenniveau, maar in veel opzichten net zo complex, is de belangrijke discussie over de definitie en afbakening van instandhoudingseenheden als belangrijke elementen van intraspecifieke diversiteit die in instandhoudingsacties moeten worden overwogen (Ryder, 1986; Waples, 1991; Dizon et al ., 1992; Moritz, 1994; Vogler and de Salle, 1994; Crandall et al., 2000; Fraser and Bernatchez, 2001; Avise, 2004; Funk et al., 2012) (Figuur 4). Gezien de voorzichtige benadering van het herkennen van soorten uit genomisch bewijs dat we hierboven pleiten (en zie ook Oliver et al., 2015; Singhal et al., 2018), is het onvermijdelijk dat sommige taxonomische soorten nog steeds lange geïsoleerde populaties binnen hen zullen bevatten. Concepten zoals Evolutionarily Significant Units (ESUs) of Designatable Units (dus) zijn voorgesteld als aanvulling op bestaande taxonomie (Ryder, 1986; Waples, 1991; Moritz, 1994; Mee et al., 2015). Net als bij soorten variëren de interpretaties en criteria, maar de gemeenschappelijke bedoeling is om belangrijke elementen van intraspecifieke diversiteit te erkennen voor instandhoudingsacties (Funk et al., 2012). Moritz (1994, 2002) pleitte voor een simplistische maar praktische moleculaire benadering van het identificeren van onafhankelijk evoluerende segmenten binnen soorten, de “historische” as van diversiteit; terwijl anderen meer nadruk leggen op de “adaptieve” as om functionele diversiteit op te nemen (Ryder, 1986; Crandall et al., 2000; Fraser and Bernatchez, 2001; Waples et al., 2001; Mee et al., 2015) (Figuur 4). Ongeacht de relatieve verdiensten van deze alternatieven, is het duidelijk dat de technische en analytische instrumenten van populatiegenomica nu krachtig beide componenten van intraspecifieke diversiteit kunnen identificeren (Funk et al., 2012; Hoban et al., 2013; Whitlock, 2014; Yang and Rannala, 2014).
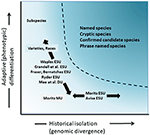
Figuur 4. Scheiding van genetische diversiteit in twee componenten: adaptieve variatie die ontstaat door natuurlijke selectie, en geanalyseerd door analyse van fenotypen en historische isolatie of neutrale divergentie beoordeeld door genomische divergentie. Het donkerder gearceerde gebied boven de curve geeft aan onder welke omstandigheden populaties in de meeste Concepten waarschijnlijk als afzonderlijke soorten worden beschouwd. Verschillende concepten en infra-specifieke taxonomische categorieën worden getoond met betrekking tot de twee assen van genetische diversiteit (na Moritz, 2002).
Dit leidt tot de vraag: Waarom noem je ESUs niet gewoon als ondersoort? Ondersoorten zijn traditioneel gedefinieerd en gezien door zowel botanici als zoölogen als fenotypisch verschillende, allopatrische groepen populaties die in elkaar kunnen intergraden op geografische grenzen (Mayr, 1942; Grant, 1981). Als zodanig weerspiegelen ze potentieel adaptieve divergentie zonder noodzakelijkerwijs historisch geïsoleerd te zijn. Ondersoorten worden veel gebruikt in planten, met geografische en ecologische verschillen de primaire manier waarop ze worden onderscheiden (Hamilton en Reichard, 1992). Voor zover ondersoorten lokale adaptatie vertonen, met of zonder voortdurende genenstroom, stellen wij voor dat fenotypisch gedefinieerde ondersoorten erkend blijven in instandhouding, en niet worden samengevoegd met ESUs, althans niet ESUs zoals gedefinieerd als historisch geïsoleerde populaties.op dezelfde manier als populatiegenomica een veel grotere precisie biedt bij de identificatie van ESUs door gebruik te maken van zowel neutrale als adaptieve loci, biedt het ook een middel om grote aantallen neutrale loci te gebruiken om Managementeenheden (mu ‘ s) gemakkelijker af te bakenen als demografisch onafhankelijke eenheden die worden gekenmerkt door een beperkte genstroom (Funk et al., 2012). Terwijl het grootste deel van onze focus hier is op ESU ’s en analoge eenheden zoals DUs die kunnen worden erkend als de belangrijkste intra-specifieke eenheden voor behoud en zijn geaccepteerd voor bescherming onder sommige rechtsgebieden, mu’ s zijn ook geïdentificeerd die demografisch onafhankelijke groepen van populaties vertegenwoordigen (zie Palsbøll et al., 2007) die kunnen worden beheerd om de grotere ESU te behouden (Moritz, 1994; Funk et al., 2012). Beheerseenheden kunnen van belang zijn voor het toezicht op en het beheer van populaties van soorten en worden ook wel “bestanden” genoemd, met name van belang voor het visserijbeheer (Avise, 2004; Laikre et al., 2005). De definitie van MUs door Moritz (1994) als “populaties met significante divergentie van allelfrequenties op nucleaire of mitochondriale loci, ongeacht de fylogenetische onderscheidend vermogen van de allelen” wordt algemeen aanvaard, hoewel, zoals aangegeven door Palsbøll et al. (2007), demografische connectiviteit is de belangrijkste factor niet het niveau van historische gen flow.het afbakenen van instandhoudingseenheden als belangrijke elementen van intraspecifieke diversiteit heeft ook belangrijke implicaties voor de genetische redding van kleine inteeltpopulaties door herstel van de genstroom of door vergroting en het wordt steeds duidelijker dat genetische redding breder moet worden overwogen om een toename van de uitsterving van de populatie te voorkomen (Love Stowell et al., 2017; Ralls et al., 2018). Het werd voorgesteld door Moritz (1999) dat het mengen van MUs maar niet ESUs kan worden beschouwd als een geschikte strategie voor genetische redding. Zoals benadrukt door Frankham et al. (2012) het mengen van taxonomisch erkende soorten zou over het algemeen niet aanvaardbaar worden geacht door managers, in welk geval taxonomische oversplitting de mogelijkheden zou kunnen beperken om kleine inteeltpopulaties te redden. Wij stellen voor dat hoewel een meer rigoureuze benadering van het afbakenen van intraspecifieke eenheden zoals MUs dit probleem zou kunnen helpen verlichten, het ook duidelijk is dat genomische schatting van migratiecijfers en van adaptieve divergentie het potentieel heeft om veel robuustere begeleiding te bieden, niet alleen bij het definiëren van soortgrenzen en intraspecifieke variatie, maar ook bij het ontwerpen van genetische reddingsstrategieën.
Fylogenomica en conservering
Genomica gecombineerd met fylogenetische methoden kunnen gemakkelijk relaties en evolutionaire afstand tussen taxa oplossen, evenals het afbakenen van soorten. Dit kan het best worden bereikt met behulp van species tree methoden, in plaats van concatenation, als de voormalige rekening voor Gen boom heterogeniteit en te hoge inflatie van tip lengtes als gevolg van voorouderlijk polymorfisme te voorkomen (Edwards et al., 2016; Ogilvie et al., 2016). Terwijl massale aantallen loci zijn vaak niet essentieel, met inbegrip van meer loci vaak verbetert de resolutie van zowel relaties en tak lengtes (bijv., Blom et al., 2016). Het belang voor het behoud is tweeledig. Ten eerste, fylogenetische beoordelingen mogelijk overweging van de onderscheidbaarheid van soorten of ESUs, niet alleen of ze worden beschreven. Dus, de rand programma (Isaac et al., 2007) pleit voor weging van soorten voor behoud op basis van hoeveel unieke evolutionaire geschiedenis ze vertegenwoordigen, in aanvulling op de gebruikelijke IUCN criteria. Ten tweede kunnen taklengten zelf biodiversiteitswaarden vertegenwoordigen, onafhankelijk van taxonomie (Faith, 1992), die waarde heeft aangetoond bij het prioriteren van gebieden voor behoud (Laity et al., 2015; Rosauer et al., 2016, 2018). In deze Betekenis, maken het goedkeuren van fylogenetic metrics gebruikend maatregelen zoals fylogenetic diversiteit en fylogenetic endemism het mogelijk om gebieden van hoge evolutionaire diversiteit te benadrukken die voor bescherming kunnen worden gericht, en hoge prioriteit in om het even welk planningsproces voor verbeterd Biodiversiteitsbehoud worden gegeven. Deze aanpak kan de bescherming van genetische diversiteit en evolutionaire processen voor diverse groepen organismen over een reeks geografische schalen (Laity et al., 2015) het maken van beslissingen over gebieden om robuuster te prioriteren aan de grillen van hoe genomic bewijsmateriaal naar erkenning van species of intra specifieke eenheden wordt vertaald.
de reële wereld—wettelijke definities
beleidskaders en wetgevingskaders voor het behoud van biodiversiteit variëren van internationale verdragen en strategieën, via nationale strategieën en wetgeving, en nationale of regionale wetgeving en strategieën binnen landen. De belangrijkste internationale instrumenten inzake biologische behoud zijn het Verdrag inzake Biologische Diversiteit geïmplementeerd door middel van de Overeenkomst van de Partijen (COP), de World Heritage Convention, de Mondiale Biodiversiteit Strategie, de Montreal Proces voor het Behoud van Bossen, de Ramsar Convention on Wetlands, de Conventie over Trekkende Soorten, de Convention on International Trade in Endangered Species (CITES), de International Union for Conservation of Nature (IUCN) op de rode lijst voor bedreigde soorten, en de United Nations Framework convention on climate change. Al deze handvesten belichamen het principe van het behoud van soorten en genetische diversiteit als een fundamenteel doel van het behoud van de biodiversiteit, waarbij soorten De ruim gedefinieerde eenheid voor bescherming zijn. De definities van soorten in wetgeving en internationale richtlijnen zijn echter zeer verschillend. Zij omvatten taxonomische erkenning van soorten en intraspecifieke taxa, maar kunnen ook genetische/evolutionaire concepten omvatten die vaak onder de wettelijke definitie van een “soort” vallen (Tabel 1).

Tabel 1. Wereldwijde wetgeving en richtlijnen die “soorten” definiëren om in de lijst te worden opgenomen en te worden beschermd.
gezien de brede waaier van infra-specifieke niveaus die kunnen worden beschermd (Tabel 1), moeten de wetgeving en de bijbehorende procedures voor het opnemen van landen het behoud van genetische diversiteit vergemakkelijken. Zo kan de definitie van een “soort”, wanneer men kijkt naar wetgeving over de hele wereld (Tabel 1), omvatten:: soorten, ondersoorten, variëteiten en geografisch en/of genetisch verschillende populaties, terwijl onder de Zuid-Afrikaanse wetgeving ook cultivars, geografische rassen, stammen en hybriden kunnen vallen. In de meeste wetgeving over de hele wereld omvat een “soort” dus entiteiten die verschillende infra-specifieke niveaus weerspiegelen die allemaal deel uitmaken van een bredere soortendistributie en in verschillende mate het populatie-soortencontinuüm weerspiegelen. Dit komt overeen met de onvermijdelijke moeilijkheid waarmee taxonomisten, evolutionaire biologen en conservatiegenetici worden geconfronteerd bij het vinden van een meer consistente en uniforme manier om eenheden binnen dit continuüm te specificeren.
sommigen beweren dat bezorgdheid over de beleidsimplicaties van soorten en infra-specifieke onzekerheid misplaatst is (Hey et al.(2003), maar wij zijn nog steeds van mening dat wetgeving weliswaar een mechanisme kan bieden voor de bescherming van genetische diversiteit op een aantal niveaus, maar dat het aan biologen is om meer duidelijkheid en consistentie te verschaffen over hoe deze niveaus van soort tot individuele populaties moeten worden afgebakend. Op dit moment blijven managers en wetgevers achter met een verbijsterende reeks soortconcepten en infra-specifieke eenheden die door wetenschappers worden geleverd bij het afbakenen van eenheden in het populatie-soortencontinuüm. Zo beschouwen alle rechtsgebieden soorten die door taxonomisten worden erkend als belangrijke eenheden voor instandhouding (Tabel 1), maar er is nog steeds een breed debat onder taxonomisten over soortconcepten, en hoe soorten in de praktijk worden gediagnosticeerd varieert binnen en tussen groepen. Dit heeft geleid tot een recente oproep voor een meer gereguleerde benadering van taxonomie (Garnett and Christidis, 2017), waarop anderen hebben tegengegaan dat regulering van taxonomie is niet het antwoord (Thomson et al., 2018). Hoewel we het eens zijn met een aantal van de punten van Thomson et al. (2018) we stellen voor dat het nogal naïef is om voor te stellen dat de wetgeving zelf moet worden gewijzigd om expliciet te verwijzen naar het specifieke taxonconcept dat wordt geïmpliceerd door een naam. Deze en eerdere debatten benadrukken de noodzaak voor evolutionaire biologen en taxonomisten om het voortouw te nemen in het waarborgen van duidelijkheid (transparantie) in hoe zij genomisch en ander bewijs vertalen naar afbakening van soorten of intraspecifieke eenheden, en hoe deze taxa zich verhouden tot relevante wetgeving voor behoud en bescherming.
vertaling naar het Management
zoals hierboven besproken, erkennen alle wetgeving en richtlijnen het soortenniveau als de fundamentele eenheid van instandhouding, maar voorzien zij ook in de erkenning van verschillende infra-specifieke eenheden. Velen hebben ook een impliciete eis voor de formele naamgeving en taxonomische beschrijving van soorten wanneer overwogen voor behoud prioritering en formele bescherming. In het kader van de IUCN Red Listing Guidelines wordt de lijst van niet nader genoemde soorten bijvoorbeeld afgeraden en wordt vereist dat de nieuwe soortbeschrijving wordt gepubliceerd binnen vier jaar nadat de soort op de IUCN Red List is opgenomen of dat de nieuwe soortbeschrijving wordt geschrapt. Dit benadrukt het belang van taxonomische naamgeving en beschrijving van entiteiten, op zowel soorten als infra-specifieke niveaus, de beperkte Taxonomische informatie voor vele planten – en diergroepen (Mace, 2004; Oliver and Lee, 2010; Wilson, 2017), en de noodzaak om taxonomische inspanningen te versnellen, met name in biodiversiteitsrijke gebieden (Riedel et al., 2013; Wege et al., 2015; Wilson, 2017).
taxonomisch benoemde soorten en intraspecifieke taxonomische categorieën, zoals ondersoorten en variëteiten, bieden over het algemeen de basiseenheden voor de formele notering, prioritering van de instandhouding en bescherming, maar in sommige rechtsgebieden worden andere categorieën van intraspecifieke instandhoudingseenheden formeel erkend. Deze worden over het algemeen geà dentificeerd gebruikend gecombineerde genomic en fenotypic gegevens. Bijvoorbeeld, de USA Endangered Species Act herkent verschillende Populatiesegmenten (DPSS) die breed zijn gebaseerd op ESUs (Waples, 1991), terwijl de Canadian Species at Risk Act DUs erkent. Een operationele interpretatie van” verschillende populaties ” onder de Canadese Species at Risk Act, dus gericht op het identificeren van discrete sets van populaties die belangrijke componenten van de evolutionaire erfenis van soorten vertegenwoordigen en die waarschijnlijk niet worden vervangen door natuurlijke verspreiding (Mee et al., 2015).
zowel DUs als DPSs overlappen conceptueel met ESUs (Waples et al., 2013) en zijn het meest toegepast op Noord-Amerikaanse vissen, met inbegrip van Noordwest-Amerikaanse zalmachtigen (Waples, 1995) en Canadese coregoniden (Mee et al., 2015) interessant zijn DPSs in de VS alleen van toepassing op gewervelde dieren, terwijl in Canada DUs kan worden toegepast op elk organisme dat kan worden vermeld onder de wet (zie Waples et al., 2013), met DUs momenteel opgenomen voor geleedpotigen, weekdieren en planten. In Australië, net als Canada, kunnen verschillende populaties worden vermeld voor elk organisme erkend onder de Environment Protection and Biodiversity Conservation Act 1999 (EPBC Act), hoewel ze niet een specifiek genoemde categorie, zoals DUSs en DPSs. Tot op heden zijn ze echter alleen erkend voor gewervelde dieren. In de VS zijn 127 gewervelde DPS ‘ s opgenomen voor behoud (https://ecos.fws.gov/ecp/); In Canada zijn dus 70 dus opgenomen voor alle organismen (http://www.registrelep-sararegistry.gc.ca/species/schedules_e.cfm?id=1); en in Australië zijn populaties van 7 gewervelde soorten waaronder zoogdieren, vissen en vogels krachtens de EPBC-wet erkend als met een specifieke instandhoudingsstatus (http://www.environment.gov.au/cgi-bin/sprat/public/publicthreatenedlist.pl). Terwijl beoordelingen van deze populaties in de VS en Canada over het algemeen gebruik maken van genetische informatie (Waples et al., 2013), in Australië zijn ze meestal gebaseerd op geografische en jurisdictionele identificatie, hoewel in sommige van deze gevallen zijn ze ondersteund door analyse van genetische differentiatie voor zoogdieren (Firestone et al., 1999; Armstrong, 2006) en fish (Colgan and Paxton, 1997; Ahonen et al., 2009).
de lijst van infra-specifieke instandhoudingseenheden voor planten en ongewervelde dieren in Canada (DUs) maar niet in de VS of Australië belicht een ander probleem dat niet specifiek verband houdt met de afbakening van instandhoudingseenheden, maar meer in het algemeen met significante inconsistenties in de prioritering van Biodiversiteitsbehoud; iconische gewervelde dieren zoals vogels, zoogdieren en vissen zijn een aandachtspunt in veel rechtsgebieden, terwijl andere organismen dat niet zijn. Deze preferentiële lijst van populaties van bepaalde organismen weerspiegelt een reeks kwesties die minder te maken hebben met de wetenschap van het afbakenen van beschermingseenheden en meer te maken hebben met de lijst van populaties op basis van publiek bewustzijn, politieke opportuniteit en de charismatische aard van het organisme dat moet worden vermeld (zie Waples et al., 2013).hoewel in Australië sommige populaties van bekende zoogdieren, vogels en vissen beschermd zijn op grond van de EPBC-wet, is er nu een significante hoeveelheid informatie op basis van moleculaire studies bij reptielen, ongewervelde dieren en planten, waar populaties kunnen worden herkend als ESUs of MUs, en veel van deze populaties zijn van belang voor het behoud gezien hun beperkte geografische spreiding, kleine populatie, vervalsnelheid en gevoeligheid voor bedreigende processen. Bijvoorbeeld, opvallende voorbeelden van zeer gestructureerde populaties en sterk uiteenlopende geslachten binnen soorten die gemakkelijk zouden voldoen aan de eis van een ESU kunnen worden gevonden in een reeks van laag verspreid gewervelde dieren uit de tropen (Figuur 2) en elders. Ook in Australische ongewervelde dieren, zoals amfipoden en miljoenpoten, ondersteunen significante genetische structuur en fylogeografische patronen de afbakening van discrete populatieclusters als ESUs die geografisch ongelijksoortig zijn, met smalle geografische bereiken, beperkt tot specifieke habitats en vertegenwoordigd door slechts een paar populaties (Finston et al., 2007; Murphy et al., 2013; Nistelberger et al., 2014a). Vaak zijn deze korte-afstand endemische taxa van zorg voor het behoud (Harvey et al., 2011) en zijn mede-gevestigd in endemism hotspots (e.g., Oliver et al., 2017), die zelf hoge prioriteiten voor multi-taxon behoud aandacht (Rosauer et al., 2018), zoals de Kimberley regio, en de Stirling Range in het zuidwesten van West-Australië.in Australische planten zijn er ook een significant aantal genetische en fylogeografische populatiestudies die verschillende populatiegroepen identificeren binnen een reeks soorten van verschillende geslachten die disjuncte en historisch geïsoleerde populatiesystemen, geologische en edafische complexiteiten weerspiegelen, en die contrasterende habitats bezetten in termen van vegetatie en Klimaat (zie Broadhurst et al., 2017). Genetisch verschillende populaties zijn bijzonder duidelijk in soorten uit Zuidwest West Australië (Coates, 2000; Byrne et al., 2014) met een aantal studies die significante genetische structuur en zeer uiteenlopende geslachten in een reeks van plantengeslachten en families met inbegrip van benadrukken: Acacia (Mimosaceae; Coates, 1988), Atriplex (Amaranthaceae; Shepherd et al., 2015), Banksia (Proteaceae; Coates et al., 2015), Lambertia (Proteaceae; Byrne et al., 2002), Hakea (Proteaceae; Sampson et al., 2014), Eremophila (Scrophulariaceae; Llorens et al. Bladvlek: Verkleurd, Meestal Niet Vergald, Vaak ± Necrotisch Teken Van Een Schimmelaantasting; Millar and Byrne, 2013), en Calothamnus (Myrtaceae; Nistelberger et al., 2014b). In veel gevallen worden deze populaties erkend in het behoud van de planning door de staat overheidsinstellingen en rekening gehouden in milieueffectrapportages (zie Shepherd et al., 2015), ook al staan ze niet op de lijst voor bescherming onder wetgeving. We erkennen echter dat de formele erkenning van deze eenheden hun instandhouding op lange termijn zou verbeteren.
Deze Australische voorbeelden benadrukken de toenemende behoefte om deze laag van genetische diversiteit onder het soortenniveau te herkennen. De uitdaging is ervoor te zorgen dat zeer gestructureerde populaties en sterk uiteenlopende geslachten die van belang zijn voor het behoud, worden beschermd tegen concurrerende eisen voor aandacht voor het behoud. Naast iconische soorten is deze uitdaging ook wereldwijd duidelijk, met uitzondering van Canada. Het niet beschermen van duidelijk afgebakende instandhoudingseenheden zal waarschijnlijk het risico op cryptische uitsterving en verlies van significante genetische diversiteit verhogen, wat in veel gevallen kan leiden tot afstammelingen en historisch geïsoleerde populaties die al miljoenen jaren bestaan binnen soorten (Moritz, 2002).
conclusies en aanbevelingen
de lijst van soorten en verschillende infra-specifieke categorieën in het kader van de wetgeving inzake bedreigde soorten of in het kader van internationale organisaties zoals IUCN, duidt op een verbintenis om de genetische diversiteit in het gehele soortenpopulatiecontinuüm te beschermen. Toch richten de huidige benaderingen van het behoud van biodiversiteit zich grotendeels op het soorteneinde van dit continuüm, met minder aandacht voor genetische diversiteit en infra-specifieke variatie. Deze meer beperkte focus op intra-species niveau genetische variatie suggereert dat, terwijl instandhoudingsbeheerders steeds meer de waarde erkennen van expliciet rekening houden met genetische informatie in beheer acties en prioritering, ze vaak gebrek aan bruikbare informatie van biodiversiteitswetenschappers (Waples et al., 2008; Laikre et al., 2010; Cook and Sgrò, 2017; Taylor et al., 2017). Deze kloof tussen wetenschap en instandhoudingspraktijk belemmert een effectievere globale aanpak van het behoud en het beheer van genetische diversiteit.
wij stellen voor dat het gebrek aan duidelijkheid van biodiversiteitswetenschappers over instandhoudingseenheden grotendeels gebaseerd is op inconsistenties op twee niveaus. De eerste betreft de concepten van meerdere soorten en de verschillende diagnostische methoden die worden gebruikt om soorten af te bakenen die leiden tot inconsistentie tussen taxonomische groepen. De tweede heeft betrekking op een gebrek aan consistentie en overeenstemming tussen taxonomisten, evolutionaire biologen en conservatiebiologen over de behandeling van infra-specifieke taxa en andere entiteiten zoals ESUs, DUs en MUs.
hoewel wij van mening zijn dat de wetgeving vaak al voldoende flexibel is wat betreft de definitie van taxa (of “soorten”), stellen wij dat het noodzakelijk is om systematisch het continuüm van populatiesoorten aan te pakken en duidelijker eenheden voor instandhouding te definiëren, waarbij begrippen als Ege ‘ s meer erkenning krijgen en hoe deze zich verhouden tot relevante wetgeving. We benadrukken dat deze kwestie ongetwijfeld kritischer zal worden gezien de kracht van genomica om differentiatie op steeds fijnere schaal te detecteren en het toenemende gebruik ervan om soorten en infra-specifieke entiteiten af te bakenen.
uit de voorgaande analyse geven we de volgende aanbevelingen:
(i) genomische gegevens moeten conservatief worden geïnterpreteerd bij het afbakenen van soorten, gebruikmakend van een integratieve taxonomische benadering die doorgaans bewijs omvat dat verder gaat dan alleen de genetische gegevens.
(ii) Waar genetici identificeren van meerdere lijnen of verschillende populaties binnen een soort, en deze niet garanderen hoogte te species niveau en zijn niet taxonomisch erkend, dat er een sterkere impuls en een meer uniforme aanpak te identificeren duidelijk gemotiveerd intraspecific eenheden voor de praktische beoordeling van de staat van instandhouding of impact assessments, en dat deze expliciet worden erkend als eenheden voor instandhouding in het kader van de wetgeving relevant voor deze jurisdictie.
iii) de IUCN zal, via haar gespecialiseerde groep Conservation Genetics en haar regionale subgroepen, richtsnoeren ontwikkelen voor de erkenning en etikettering van intraspecifieke eenheden die aansluiten bij de beste wetenschappelijke praktijk.
(iv) de meer genuanceerde kijk die uit de evolutionaire Genomica van soorten en het continuüm van populaties tot soorten naar voren komt, moet door wetenschappers effectiever worden gecommuniceerd aan instandhoudingsbeheerders en beleidsmakers.meer in het algemeen sluiten we ons aan bij de aanbevelingen die onlangs zijn gedaan in een aantal reviews (Frankham, 2010; Santamaría and Méndez, 2012; Hoban et al., 2013; Moritz and Potter, 2013; Laity et al., 2015; Pierson et al., 2016; Cook and Sgrò, 2017; Taylor et al., 2017) die de noodzaak benadrukken voor wetenschappers om de communicatie te verbeteren en het bewustzijn te verhogen met beleidsmakers, overheidsinstanties, niet-gouvernementele organisaties, en beoefenaars dat de resultaten van moleculaire systematiek, moleculaire taxonomie, fylogeografie, en populatie genetisch onderzoek kritische informatie over het behoud van genetische diversiteit en het beheer van evolutionaire processen kan bieden.
Auteursbijdragen
DC, MB en CM droegen allemaal bij aan de voorbereiding en herziening van het manuscript.
belangenconflict verklaring
De auteurs verklaren dat het onderzoek werd uitgevoerd zonder enige commerciële of financiële relatie die als een potentieel belangenconflict kon worden opgevat.
Dankbetuigingen
Dit project werd gesteund door de Australian Research Council (ARC Linkage Grant No. LP120200063), de Australian National University en het Western Australian Department of Biodiversity, Conservation, and Attractions.
Afonso Silva, A. C., Bragg, J. G., Potter, S., Fernandes, C., Coelho, M. M., en Moritz, C. (2017a). Tropische specialist versus klimaat generalist: diversificatie en demografische geschiedenis van zustersoorten van Carlia skinks uit Noordwest Australië. Mol. Ecol. 26:4045–4058. doi: 10.1111 / mec.14185
CrossRef Full Text / Google Scholar
Afonso Silva, A. C., Santos, N., Ogilvie, H. A., and Moritz, C. (2017b). Validatie en beschrijving van twee nieuwe noordwestelijke Australische Rainbow skinks met multispecies coalescente methoden en morfologie. PeerJ 5: e3724. doi: 10.7717/peerj.3724
PubMed Abstract / CrossRef Full Text / Google Scholar
Ahonen, H., Harcourt, R. G., and Stow, A. J. (2009). Nucleair en mitochondriaal DNA onthult isolatie van in gevaar gebrachte grijze verpleegsterhaai populaties (Carcharias taurus). Mol. Ecol. 18, 4409–4421. doi: 10.1111 / j. 1365-294X. 2009. 04377.x
PubMed Abstract / CrossRef Full Text / Google Scholar
Armstrong, K. N. (2006). Fylogeografische structuur in Rhinonicteris aurantia (Chiroptera: Hipposideridae): implicaties voor het behoud. Acta Chiropterologica 8, 63-81. doi: 10.3161/1733-5329(2006)82.0.CO
CrossRef Full Text/Google Scholar
Arnold, M. L., and Kunte, K. (2017). Adaptieve genetische uitwisseling: een verwarde geschiedenis van vermenging en evolutionaire innovatie. Trends Ecol. Evol. 32, 601–611. doi: 10.1016 / j.boom.2017.05.007
PubMed Abstract / CrossRef Full Text / Google Scholar
Avise, J. C. (2004). Moleculaire merkers, natuurlijke geschiedenis en evolutie. 2nd Edn. Sunderland, MA; Oxford: Oxford University Press.
Google Scholar
Avise, J. C., Arnold, J., Ball, R. M., Bermingham, E., Lamb, T., Neigel, J. E., et al. (1987). Intraspecifieke fylogeografie: de mitochondriale DNA-brug tussen populatiegenetica en systematiek. Anne. Rev.Ecol. Sys. 18, 489–522. doi: 10.1146 / annurev. es. 18. 110187. 002421
CrossRef Full Text / Google Scholar
Bickford, D., Lohman, D. J., Sodhi, N. S, Ng, P. K., Meier, R., Winker, K., et al. (2007). Cryptische soorten als venster op diversiteit en behoud. Trends Ecol. Evol. 22, 148–155. doi: 10.1016 / j.boom.2006.11.004
PubMed Abstract / CrossRef Full Text / Google Scholar
Blom, M. P. K., Bragg, J. G., Potter, S., and Moritz, C. (2016). Accounting for uncertainty in gene tree estimation: summary-coalescent species tree inference in a challenging radiation of Australian hagedissen. Syst. Biol. 66, 352–366. doi: 10.1093 / sysbio / syw089
PubMed Abstract / CrossRef Full Text / Google Scholar
Broadhurst, L., Breed, M., Lowe, A., Bragg, J., Catullo, R., Coates, D. J., et al. (2017). Genetische diversiteit en structuur van de Australische flora. Duiker. Distrib. 23, 41–52. doi: 10.1111 / ddi.12505
CrossRef Full Text / Google Scholar
Byrne, M., Coates, D. J., Forest, F., Hopper, S. D., Krauss, S. L., Sniderman, J. M. K., et al. (2014). “A diverse flora-species and genetic relationships,” in Plant Life on the Sandplains in Southwest Australia, a Global Biodiversity Hotspot, editors H. Lambers (Crawley, WA: University of Western Australia Publishing), 81-99.
PubMed Abstract
Byrne, M., Macdonald, B., and Coates, D. J. (2002). Phylogeografische patronen en hoge cpDNA diversiteit binnen de Acacia acuminata (Leguminosae:Mimosoideae). complex in West-Australië. J. Evol. Biol. 15, 576–587. doi: 10.1046 / j. 1420-9101. 2002. 00429.x
CrossRef Full Text / Google Scholar
Carstens, B. C., Pelletier, T. A., Reid, N. M., and Satler, J. D. (2013). Hoe te falen bij de afbakening van soorten. Mol. Ecol. 22, 4369–4383. doi: 10.1111 / mec.12413
PubMed Abstract / CrossRef Full Text/Google Scholar
Coates, D. J. (1988). Genetische diversiteit en populatiegenstructuur in de zeldzame Chitterende graswal, Acacia anomala Court. Aust. J. Bot. 36:273–286. doi: 10.1071 / BT9880273
CrossRef Full Text / Google Scholar
Coates, D. J. (2000). Het definiëren van instandhoudingseenheden in een rijke en gefragmenteerde flora: implicaties voor het beheer van genetische hulpbronnen en evolutionaire processen in Zuidwest-Australië planten. Aust. J. Bot. 48, 329–339. doi: 10.1071 / BT99018
CrossRef Full Text / Google Scholar
Coates, D. J., McArthur, S. L., and Byrne, M. (2015). Significant verlies van genetische diversiteit als gevolg van pathogeen-gedreven uitsterving in de zeldzame endemische Banksia brownii (Proteaceae). Biol. Conserv. 192, 353–360. doi: 10.1016 / j.biocon.2015.10.013
CrossRef Full Text / Google Scholar
Colgan, D. J., and Paxton, J. R. (1997). Biochemische genetica en herkenning van een westelijk bestand van de gewone edelvis, Rexea solandri (Scombroidea: Gempylidae)., in Australië. Mar. Freshw. Res. 48, 103-118. doi: 10.1071 / MF96048
CrossRef Full Text/Google Scholar
Convention on Biological Diversity (2007). Online beschikbaar op: https://www.cbd.int/doc/meetings/cop-bureau/cop-bur-2007/cop-bur-2007-10-14-en.pdf
Cook, C. N., and Sgrò, C. M. (2017). Het afstemmen van wetenschap en beleid om evolutionair verlicht behoud te bereiken. Conserv. Biol. 31, 501–512. doi: 10.1111 / cobi.12863
PubMed Abstract / CrossRef Full Text / Google Scholar
Cook, L. G., Edwards, R. D., Crisp, M. D., and Hardy, N. B. (2010). Moet morfologie altijd nodig zijn voor nieuwe soortbeschrijvingen? Inverteer. Sys. 24, 322–332. doi: 10.1071/IS10011
CrossRef Full Text/Google Scholar
Cracraft, J. (1983). Soortconcepten en speciatieanalyse. Curr. Ornithol. 1, 159–187. doi: 10.1007/978-1-4615-6781-3_6
CrossRef Full Text / Google Scholar
Crandall, K. A., Bininda-Emonds, O. R., Mace, G. M., and Wayne, R. K. (2000). Het overwegen van evolutionaire processen in conservatiebiologie. Trends Ecol. Evol. 15, 290–295. doi: 10.1016 / S0169-5347 (00)01876-0
PubMed Abstract | CrossRef Full Text | Google Scholar
Cutter, A. D., and Payseur, B. A. (2013). Genomic handtekeningen van selectie op gekoppelde plaatsen: het verenigen van de ongelijkheid tussen soorten. Nat. Eerwaarde Genet. 14, 262–274. doi: 10.1038 / nrg3425
PubMed Abstract / CrossRef Full Text / Google Scholar
Dayrat, B. (2005). Naar integratieve taxonomie. Biol. J. Linn. Soc. 85, 407–415. doi: 10.1111 / j. 1095-8312. 2005. 00503.x5
CrossRef Full Text / Google Scholar
De Queiroz, K. (1998). “The general lineage concept of species, species criteria, and the process of speciation,” in Forms: Species and Speciation, D. J. Howard, R S. H. Berlocher (New York, NY: Oxford University Press), 57-75.
Google Scholar
De Queiroz, K. (2007). Soortbegrippen en soortafbakening. Syst. Biol. 56, 879–886. doi: 10.1080 / 10635150701701083
PubMed Abstract | CrossRef Full Text/Google Scholar
Degnan, J. H., and Rosenberg, N. A. (2009). Gen tree discordance, fylogenetic inference en de multispecies coalescent. Trends Ecol. Evol. 24, 332–340. doi: 10.1016 / j.boom.2009.01.009
PubMed Abstract / CrossRef Full Text / Google Scholar
Devey, D. S., Bateman, R. M., Fay, M. F., and Hawkins, J. A. (2008). Vrienden of familie? Fylogenetica en soortafbakening in het controversiële Europese orchideeëngeslacht Ophrys. Anne. Robot. 101, 385–402. doi: 10.1093 / aob / mcm299
PubMed Abstract / CrossRef Full Text / Google Scholar
Dizon, A. E., Lockyer, C, Perrin, W. F., Demaster, D. P., and Sisson, J. (1992). Herdenken van het stock concept-een fylogeografische aanpak. Conserv. Biol. 6, 24–36. doi: 10.1046 / j. 1523-1739.1992.610024.X
CrossRef Full Text / Google Scholar
Doughty, P., Bourke, G., Tedeschi, L., Pratt, R., Oliver, P., Palmer, R., et al. (2018). Afbakening van soorten in de Gehyra nana (Squamata: Gekkonidae). complex: cryptische en divergente morfologische evolutie in de Australische Moessoonkring, met de beschrijving van vier nieuwe soorten. Zootaxa, 4403: 201-244. doi: 10.11646/zootaxa.4403.2.1
PubMed Abstract / CrossRef Full Text / Google Scholar
Dynesius, M., and Jansson, R. (2014). Persistentie van binnen-soorten geslachten: een verwaarloosde controle van speciatiesnelheden. Evolutie 68, 923-934. doi: 10.1111 / evo.12316
PubMed Abstract / CrossRef Full Text/Google Scholar
Edwards, S. V., Potter, S., Schmitt, C. J., Bragg, J. G., and Moritz, C. (2016). Reticulation, divergentie, and the phylogeography-phylogenetics continuumc. Proc. Natl. Acad. Sci. U. S. A. 113, 8025-8032. doi: 10.1073 / pnas.1601066113
CrossRef Full Text / Google Scholar
Faith, D. P. (1992). Conservatie-evaluatie en fylogenetische diversiteit. Biol. Nadeel. 61, 1–10. doi: 10.1016/0006-3207(92)91201-3
CrossRef Full Text / Google Scholar
Finston, T. L., Johnson, M. S., Humphreys, W. F., Eberhard, S. M., and Halse, S. A. (2007). Cryptische soortvorming in twee wijdverspreide onderaardse amfipodengeslachten weerspiegelt historische drainagepatronen in een oud landschap. Mol. Ecol. 16, 355–365. doi: 10.1111 / j. 1365-294X. 2006. 03123.x
PubMed Abstract / CrossRef Full Text/Google Scholar
Firestone, K. B., Elphinstone, M. S., Sherwin, W. B., and Houlden, B. A. (1999). Fylogeografische populatiestructuur van de tijger quolls Dasyurus maculatus (Dasyuridae: Marsupalia), een bedreigde vleesetende buideldier. Mol. Ecol. 8, 1613–1625. doi: 10.1046 / j. 1365-294x. 1999. 00745.x
CrossRef Full Text / Google Scholar
Fišer, C., Robinson, C. T., and Malard, F. (2018). Cryptische soorten als venster op de paradigmaverschuiving van het soortenconcept. Mol. Ecol. 27, 613–635. doi: 10.1111 / mec.14486
PubMed Abstract / CrossRef Full Text/Google Scholar
Frankham, R. (2010). Uitdagingen en kansen van genetische benaderingen van biologische instandhouding. Biol. Conserv. 143,1919–1927. doi: 10.1016 / j.biocon.2010.05.011
CrossRef Full Text / Google Scholar
Frankham, R., Ballou, J. D., Dudash, M. R., Eldridge, M. D., Fenster, C. B., Lacy, R. C., et al. (2012). Implicaties van verschillende soortenconcepten voor het behoud van de biodiversiteit. Biol. Conserv. 153, 25–31. doi: 10.1016 / j.biocon.2012.04.034
CrossRef Full Text / Google Scholar
Fraser, D. J., and Bernatchez, L. (2001). Adaptive evolutionary conservation: naar een uniform concept voor het definiëren van conservatie-eenheden. Mol. Ecol. 10, 2741–2752. doi: 10.1046 / j. 1365-294X.2001.t01-1-01411.x
PubMed Abstract / CrossRef Full Text/Google Scholar
Funk, W. C., Mckay, J. K., Hohenlohe, P. A., and Allendorf, F. W. (2012). Gebruik maken van genomica voor het afbakenen van conservatie-eenheden. Trends Ecol. Evol. 27, 489–496. doi: 10.1016 / j.boom.2012.05.012
PubMed Abstract / CrossRef Full Text / Google Scholar
Garnett, S. T., and Christidis, L. (2017). Taxonomie anarchie belemmert behoud. Natuur 546, 25-27. doi: 10.1038 / 546025a
PubMed Abstract / CrossRef Full Text / Google Scholar
Gippoliti, S., Cotterill, F. P. D., Zinner, D., and Groves, C. P. (2018). Impact van taxonomische inertie voor het behoud van de diversiteit van de Afrikaanse hoefdieren: overzicht. Biol. Rev. 93, 115-130. doi: 10.1111/brv.12335
PubMed Abstract / CrossRef Full Text/Google Scholar
Grant, V. (1981). Plantenspeciatie. New York, NY: Columbia University Press.
PubMed Abstract/Google Scholar
Hamilton, C. W., and Reichard, S. H. (1992). Huidige praktijk in het gebruik van ondersoorten, variëteiten en forma in de classificatie van wilde planten. Taxon 41, 485-498. doi: 10.2307 / 1222819
CrossRef Full Text / Google Scholar
Harvey, M. S., Rix, M. G., Framenau, V. W., Hamilton, Z. R., Johnson, M. S., Teale, R. J., et al. (2011). De onschuldigen beschermen: het bestuderen van endemische taxa op korte afstand verbetert de resultaten van het behoud. Inverteer. Syst. 25, 1–10. doi: 10.1071/IS11011
CrossRef Full Text / Google Scholar
Hey, J., Waples, R. S., Arnold, M. L., Butlin, R. K., and Harrison, R. G. (2003). Het begrijpen en confronteren van soorten onzekerheid in biologie en behoud. Trends Ecol. Evol. 18, 597–603. doi: 10.1016 / j.boom.2003.08.014
CrossRef Full Text / Google Scholar
Hoban, S. M., Hauffe, H. C., Perez-Espona, S., Arntzen, J. W., Bertorelle, G., Bryja, J., et al. (2013). Genetische diversiteit op de voorgrond plaatsen in het instandhoudingsbeleid en het beheer. Conserv. Genet. Resour. 5, 593–598. doi: 10.1007 / s12686-013-9859-y
CrossRef Full Text / Google Scholar
Isaac, N. J., Mallet, J., and Mace, G. M. (2004). Taxonomische inflatie: zijn invloed op macro-economie en behoud. Trends Ecol. Evol. 19, 464–469. doi: 10.1016 / j.boom.2004.06.004
PubMed Abstract / CrossRef Full Text/Google Scholar
Isaac, N. J., Turvey, S. T., Collen, B., Waterman, C., en Baillie, J. E. (2007). Zoogdieren op de rand: behoudsprioriteiten gebaseerd op bedreiging en fylogenie. PLoS ONE 2: e296. doi: 10.1371 / journal.pone.0000296
PubMed Abstract / CrossRef Full Text / Google Scholar
Jones, M. R., and Good, J. M. (2016). Gerichte vangst in evolutionaire en ecologische genomica. Mol. Ecol. 25, 185–202. doi: 10.1111 / mec.13304
PubMed Abstract / CrossRef Full Text / Google Scholar
Jorger, K. M., and Schrödl, M. (2013). Hoe een cryptische soort te beschrijven? Praktische uitdagingen van moleculaire taxonomie. Voorkant. Zool. 10:59. doi: 10.1186/1742-9994-10-59
PubMed Abstract / CrossRef Full Text/Google Scholar
Laikre, L., Allendorf, F., Aroner, L. C., Baker, S., Gregovich, D. P., Hansen, M. M., et al. (2010). Verwaarlozing van genetische diversiteit bij de uitvoering van het Verdrag inzake biologische diversiteit. Conserv. Biol. 24, 86–88. doi: 10.1111 / j. 1523-1739. 2009. 01425.x
PubMed Abstract / CrossRef Full Text / Google Scholar
Laikre, L., Palm, S., and Ryman, N. (2005). Genetische populatiestructuur van vissen: implicaties voor het beheer van kustgebieden. Ambio 34.11-119. doi: 10.1579/0044-7447-34.2.111
PubMed Abstract / CrossRef Full Text/Google Scholar
Laity, T., Laffan, S. W., González-Orozco, C. E., Faith, D. P., Rosauer, D. F., Byrne, M., et al. (2015). Phylodiversity to inform conservation policy: een Australisch voorbeeld. Sci. Total Environ. 534, 131–143. doi: 10.1016 / j.scitotenv.2015.04.113
PubMed Abstract / CrossRef Full Text / Google Scholar
Leache, A. D., and Oaks, J. R. (2017). Het nut van single nucleotide Polymorphism (SNP). gegevens in fylogenetica. Anne. Rev.Ecol. Syst. 48, 69–84. doi: 10.1146/annurev-ecolsys-110316-022645
CrossRef Full Text | Google Scholar
Llorens, T. M., Macdonald, B., McArthur, S., Coates, D. J., and Byrne, M. (2015). Disjunct, highly divergent genetic lineages within two rare Eremophila (Scrophulariaceae: Myoporeae). species in a biodiversity hotspot: implications for taxonomy and conservation. Biol. J. Linn. Soc. 177, 96–111. doi: 10.1111/boj.12228
CrossRef Full Text | Google Scholar
Love Stowell, S. M., Pinzone, C. A., and Martin, A. P. (2017). Het wegnemen van belemmeringen voor actieve interventies voor genetische diversiteit. Biodivers. Conserv. 26, 1753–1765. doi: 10.1007 / s10531-017-1330-z
CrossRef Full Text / Google Scholar
Mace, G. M. (2004). De rol van taxonomie in het behoud van soorten. Philos. Transvetzuren. R. Soc. B: Biol. Sci. 359, 711–719. doi: 10.1098 / rstb.2003.1454
PubMed Abstract / CrossRef Full Text / Google Scholar
Mallet, J. (2007). Hybride specie. Nature 446, 279-283. doi: 10.1038 / nature05706
PubMed Abstract | CrossRef Full Text/Google Scholar
Mayr, E. (1942). Systematiek en de oorsprong van soorten. New York, NY: Columbia University Press.
Google Scholar
Mee, J. A., Bernatchez, L., Reist, J. D., Rogers, S. M., and Taylor, E. B. (2015). Identificatie van ontwerpbare eenheden voor intraspecifieke prioritering van de instandhouding: een hiërarchische benadering toegepast op het meer van witvis soorten complex (Coregonus spp.). Evol. Appl. 8, 423–441. doi: 10.1111 / eva.12247
PubMed Abstract | CrossRef Full Text | Google Scholar
Millar, M. A., and Byrne, M. (2013). Cryptic divergent lineages of Pultenaea pauciflora M.B. Scott (Fabaceae: Mirbelieae) exhibit different evolutionary history. Biol. J. Linn. Soc. 108, 871–881. doi: 10.1111/bij.12002
CrossRef Full Text | Google Scholar
Moritz, C. (1994). Defining evolutionarily significant units for conservation. Trends Ecol. Evol. 9, 373–375. doi: 10.1016/0169-5347(94)90057-4
CrossRef Full Text | PubMed Abstract
Moritz, C. (1999). Instandhoudingseenheden en translocaties: strategieën voor het behoud van evolutionaire processen. Hereditas 130, 217-228. doi: 10.1111 / j. 1601-5223.1999.00217.x
CrossRef Full Text/Google Scholar
Moritz, C. (2002). Strategieën ter bescherming van de biologische diversiteit en de evolutionaire processen die haar ondersteunen. Syst. Biol. 51, 238–254. doi: 10.1080 / 10635150252899752
PubMed Abstract | CrossRef Full Text/Google Scholar
Moritz, C., and Potter, S. (2013). Het belang van een evolutionair perspectief in de planning van het instandhoudingsbeleid. Mol. Ecol. 22, 5969–5971. doi: 10.1111 / mec.12565
PubMed Abstract / CrossRef Full Text/Google Scholar
Moritz, C., Pratt, R. C., Bank, S., Bourke, G., Bragg, J. G., Doughty, P., et al. (2018). Cryptische afstamming diversiteit, lichaamsgrootte divergentie, en sympatry in een species complex van Australische hagedissen (Gehyra). Evolutie 27, 54-66. doi: 10.1111 / evo.13380
CrossRef Full Text / Google Scholar
Morrison, W. R., Lohr, J. L., Duchen, P., Wilches, R., Trujillo, D., Mair, M., et al. (2009). De impact van taxonomische veranderingen op instandhouding: Doodt het, kan het redden, of is het gewoon irrelevant? Biol. Nadeel. 142, 3201–3206. doi: 10.1016 / j.biocon.2009.07.019
CrossRef Full Text / Google Scholar
Murphy, N. P., Adams, M., Guzik, M. T., and Austin, A. D. (2013). Buitengewoon micro-endemisme in Australische woestijn voorjaar amfipoden. Mol. Phylogenet. Evol. 66, 645–653. doi: 10.1016/j.ympev.2012.10.013
PubMed Abstract / CrossRef Full Text / Google Scholar
Murphy, N. P., King, R. A., and Delean, S. (2015). Soort, ESUs of populaties? Het afbakenen en beschrijven van morfologisch cryptische diversiteit in Australische woestijn voorjaar amfipoden. Inverteer. Syst. 29, 457–467. doi: 10.1071/IS14036
CrossRef Full Text / Google Scholar
Nater, A., Mattle-Greminger, M. P., Nurcahyo, A., Nowak, M. G., De Manual, M., Desai, T., et al. (2017). Morfometrisch, gedragsmatig en genomisch bewijs voor een nieuwe orang-oetan soort. Curr. Cel Biol. 27, 3487–3498. doi: 10.1016 / j.cub.2017.09.047
CrossRef Full Text / Google Scholar
Nicotra, A. B., Chong, C., Bragg, J. G., Ong, C. R., Aitken, N. C., Chuah, A., et al. (2016). Populatie en fylogenomic ontleding via genotyping-by-sequencing in Australisch Pelargonium. Mol. Ecol. 25, 2000–2014. doi: 10.1111 / mec.13584
PubMed Abstract / CrossRef Full Text / Google Scholar
Nistelberger, H., Byrne, M., Coates, D., and Roberts, J. D. (2014a). Sterke fylogeografische structuur in een duizendpoot wijst op Pleistoceen vicariantie tussen populaties op banded iron formations in semi-arid Australië. PLoS ONE 9: e0093038. doi: 10.1371 / journal.pone.0093038
PubMed Abstract / CrossRef Full Text / Google Scholar
Nistelberger, H., Gibson, N., Macdonald, B., Tapper, S.-L., and Byrne, M. (2014b). Fylogeografisch bewijs voor twee mesic refugia in een biodiversiteit hotspot. Erfelijkheid 113, 454-463. doi: 10.1038 / hdy.2014.46
PubMed Abstract / CrossRef Full Text / Google Scholar
Ogilvie, H. A., Heled, J., Xie, D., and Drummond, A. J. (2016). Computationele prestaties en statistische nauwkeurigheid van beast en vergelijkingen met andere methoden. Syst. Biol. 65, 381–396. doi: 10.1093 / sysbio / syv118
PubMed Abstract | CrossRef Full Text/Google Scholar
Oliver, P. M., Keogh, J. S., and Moritx, C. (2015). New approaches to catalogisering and understanding evolutionary diversity: a perspective from Australian herpetology. Aust. J. Zool. 62, 417–430. doi: 10.1071 / ZO14091
CrossRef Full Text / Google Scholar
Oliver, P. M., Laver, R. J., Demello Martins, F., Pratt, R. C., Hunjan, S., and Moritz, C. (2017). Een nieuwe hotspot van gewervelde endemisme en een evolutionaire refugium in tropisch Australië. Duiker. Distrib. 23, 53–66. doi: 10.1111 / ddi.12506
CrossRef Full Text / Google Scholar
Oliver, P. M., and Lee, M. S. Y. (2010). De botanische en zoölogische codes belemmeren biodiversiteitsonderzoek door publicatie van niet nader genoemde nieuwe soorten te ontmoedigen. Taxon 59, 1201-1205. doi: 10.2307 / 20773990
CrossRef Full Text / Google Scholar
Padial, J., Miralles, A., De La Riva, I., and Vences, M. (2010). De integratieve toekomst van de taxonomie. Voorkant. Zool. 7:16. doi: 10.1186/1742-9994-7-16
PubMed Abstract / CrossRef Full Text / Google Scholar
Palsbøll, P. J., Bérubé, M., and Allendorf, F. W. (2007). Identificatie van managementeenheden met behulp van genetische populatiegegevens. Trends Ecol. Evol. 22, 11–16. doi: 10.1016 / j.boom.2006.09.003
PubMed Abstract / CrossRef Full Text / Google Scholar
Patton, J. L., and Smith, M. F. (1994). Paraphyly, polyphyly, and the nature of species borders in Pocket Gophers (Genus Thomomys). Syst. Biol. 43, 11–26. doi: 10.1093 / sysbio / 43.1.11
CrossRef Full Text / Google Scholar
Pierson, J. C., Coates, D. J., Oostermeijer, J. G. B., Beissinger, S. R., Bragg, J. G., Sunnucks, P., et al. (2016). Genetische factoren in bedreigde soorten herstelplannen op drie continenten. Voorkant. Ecol. Environ. 14, 433–440. doi: 10.1002 / fee.1323
CrossRef Full Text / Google Scholar
Potter, S., Bragg, J. G., Peter, B. M., Bi, K., and Moritz, C. (2016). Phylogenomics at the tips: inferring lineages en hun demografische geschiedenis in een tropische hagedis, Carlia amax. Mol. Ecol. 25, 1367–1380. doi: 10.1111/mec.13546
PubMed Abstract / CrossRef Full Text / Google Scholar
Potter, S., Xue, A. T., Bragg, J. G., Rosauer, D. F., Roycroft, E. J., and Moritz, C. (2018). Pleistoceen klimaatveranderingen drijven diversificatie over een tropische savanne. Mol. Ecol. 27, 520–532. doi: 10.1111 / mec.14441/full
PubMed Abstract / CrossRef Full Text / Google Scholar
Rabosky, D. L., Hutchinson, M. N., Donnellan, S. C., Talaba, A. L., and Lovette, I. J. (2014). Fylogenetic disassembly of species boundaries in a wijdverspreide group of Australian skinks (Scincidae: Ctenotus). Mol. Phylogenet. Evol. 77, 71–82. doi: 10.1016/j.ympev.2014.03.026
PubMed Abstract / CrossRef Full Text / Google Scholar
Ralls, K., Ballou, J. D., Dudash, M. R., Eldridge, M. D. B., Fenster, C. B., Lacy, R. C., et al. (2018). Roep op tot een paradigmaverschuiving in het genetisch beheer van gefragmenteerde populaties. Conserv. Lett. 11, 1–6. doi: 10.1111 / conl.124126
CrossRef Full Text / Google Scholar
Rannala, B. (2015). De kunst en wetenschap van de afbakening van soorten. Curr. Zool. 61, 846–853. doi: 10.1093/czoolo/61.5.846
CrossRef Full Text/Google Scholar
Riedel, A., Sagata, K., Suhardjono, Y. R., Tänzler, R., and Balke, M. (2013). Integrative taxonomy on the fast track-towards more sustainability in biodiversity research. Voorkant. Zool. 10:15. doi: 10.1186 / 1742-9994-10-15
PubMed Abstract | CrossRef Full Text/Google Scholar
Rieseberg, L. H., and Willis, J. H. (2007). Plantenspeciatie. Wetenschap 317, 910-914. doi: 10.1126 / wetenschap.1137729
PubMed Abstract / CrossRef Full Text/Google Scholar
Rosauer, D. F., Blom, M. P. K., Bourke, G., Catalano, S., Donnellan, S., Mulder, E., et al. (2016). Fylogeografie, hotspots en conservatieprioriteiten: een voorbeeld uit de Top van Australië. Biol. Nadeel. 204, 83–93. doi: 10.1016 / j.biocon.2016.002
CrossRef Full Text / Google Scholar
Rosauer, D. F., Byrne, M., Blom, M. P. K., Coates, D. J., Donnellan, S., Doughty, P., et al. (2018). Real-world conservation planning for evolutionary diversity in the Kimberley, Australië, omzeilt onzekere taxonomie Conserv. Lett. 11: e12438. doi: 10.1111 / conl.12438
CrossRef Full Text / Google Scholar
Rosenblum, E. B., Sarver, B. A. J., Brown, J. W., Des Roches, S., Hardwick, K. M., Hether, T. D., et al. (2012). Goldilocks meets Santa Rosalia: een kortstondig speciatiemodel verklaart patronen van diversificatie over tijdsschalen. Evol. Biol. 39, 255–261. doi: 10.1007 / s11692-012-9171-x
PubMed Abstract / CrossRef Full Text / Google Scholar
Roux, C., Fraïsse, C., Romiguier, J., Anciaux, Y., Galtier, N., and Bierne, N. (2016). Licht werpen op de grijze zone van speciatie langs een continuüm van genomische divergentie. PLoS Biol. 14: e2000234. doi: 10.1371 / journal.pbio.2000234
PubMed Abstract / CrossRef Full Text/Google Scholar
Ryder, O. A. (1986). Species conservation and systematics – het dilemma van ondersoorten. Trends Ecol. Evol. 1, 9–10. doi: 10.1016/0169-5347 (86)90059-5
CrossRef Full Text | Google Scholar
Sampson, J. F., Byrne, M., Yates, C. J., Gibson, N., Thavornkanlapachai, R., Stankowski, S., et al. (2014). Hedendaagse pollengemedieerde genenimigratie weerspiegelt de historische isolatie van een zeldzame struik in een gefragmenteerd landschap. Erfelijkheid 112, 172-181. doi: 10.1038 / hdy.2013.89
PubMed Abstract / CrossRef Full Text / Google Scholar
Santamaría, L., and Méndez, P. F. (2012). Ontwikkeling van het biodiversiteitsbeleid: huidige lacunes en toekomstige behoeften. Evol. Appl. 5, 202–218. doi: 10.1111 / j. 1752-4571. 2011. 00229.x
PubMed Abstract / CrossRef Full Text/Google Scholar
Schlick-Steiner, B. C., Steiner, F. M., Seifert, B., Stauffer, C., Christian, E., and Crozier, R. H. (2010). Integratieve taxonomie: een multisource benadering van het verkennen van biodiversiteit. Annu. Eerwaarde Entomol. 55, 421–438. doi: 10.1146/annurev-ento-112408-085432
PubMed Abstract / CrossRef Full Text / Google Scholar
Seehausen, O, Takimoto, G., Roy, D., and Jokela, J. (2008). Soortomkering en biodiversiteitsdynamiek met hybridisatie in veranderende omgevingen. Mol Ecol. 17, 30–44. doi: 10.1111 / j. 1365-294X. 2007. 03529.x
PubMed Abstract / CrossRef Full Text / Google Scholar
Shepherd, K. A., Thiele, K. R., Sampson, J., Coates, D. J., and Byrne, M. (2015). Een zeldzame, nieuwe soort van Atriplex (Chenopodiaceae) bestaande uit twee genetisch verschillende maar morfologisch cryptische populaties in aride West-Australië: implicaties voor taxonomie en behoud. Aust. Syst. Robot. 28, 234–245. doi: 10.1071/SB15029
CrossRef Full Text/Google Scholar
Singhal, S., and Bi, K. (2017). Geschiedenis ruimt rommel op: de impact van tijd in het drijven van divergentie en introgressie in een tropische hechtingszone. Evolutie 71, 1888-1899. doi: 10.1111 / evo.13278
PubMed Abstract / CrossRef Full Text / Google Scholar
Singhal, S., Hoskin, C., Couper, P., Potter, S., and Moritz, C. (2018). Framework for resolving cryptic species: een case study van de hagedissen van de Australische natte tropen. Syst. Biol. . doi: 10.1093 / sysbio / syy
PubMed Abstract | CrossRef Full Text/Google Scholar
Singhal, S., and Moritz, C. (2013). Reproductieve isolatie tussen fylogeografische lijnen schalen met divergentie. Proc. Roy. Soc B 280: 20132246. doi: 10.1098 / rspb.2013.2246026
PubMed Abstract / CrossRef Full Text/Google Scholar
Steane, D. A., Potts, B. M., McLean, E., Collins, L., Prober, S. M., Stock, W. D., et al. (2015). Genoom – brede scans onthullen cryptische populatiestructuur in een droog-aangepaste Eucalyptus. Boom Genet. Genomen 11: 33. doi: 10.1007 / s11295-015-0864-z
CrossRef Full Text / Google Scholar
Struck, T. H., Feder, J. L., Bendiksby, M., Birkeland, S., Cerca, J., Gusarov, V. I., et al. (2017). Het vinden van evolutionaire processen verborgen in cryptische soorten. Trends Ecol. Evol. 33, 153–163. doi: 10.1016 / j.boom.2017.11.007
PubMed Abstract / CrossRef Full Text / Google Scholar
Sukumaran, J., and Knowles, L. L. (2017). De multispecies coalescent begrenst structuur niet soorten. Proc. Nat. Aca. Sci. U. S. A. 114, 1607-1612. doi: 10.1073 / pnas.1607921114
PubMed Abstract / CrossRef Full Text / Google Scholar
Swarts, N. D., Clements, M. A., Bower, C. C., and Miller, J. T. (2014). Het definiëren van beschermingseenheden in een complex van morfologisch vergelijkbare, seksueel misleidende, zeer bedreigde orchideeën. Biol. Nadeel. 174, 55–64. doi: 10.1016 / j.biocon.2014.03.017
CrossRef Full Text / Google Scholar
Tautz, D., Arctander, P., Minelli, A., Thomas, R. H., and Vogler, A. P. (2003). Een pleidooi voor DNA taxonomie. Trends Ecol. Evol. 18, 70–74. doi: 10.1016 / S0169-5347 (02)00041-1
CrossRef Full Text | Google Scholar
Taylor, H. R., Dussex, N., en van Heezik, Y. (2017). Het overbruggen van de genetische kloof op het gebied van natuurbehoud door belemmeringen voor de implementatie voor natuurbeschermers in kaart te brengen. Klodder. Ecol. Conserv. 10, 231–242. doi: 10.1016/j.gecco.2017.04.001
CrossRef Full Text / Google Scholar
Thomson, S. A., Pyle, R. L., Ahyong, S. T., Alonso-Zarazaga, M., Ammirati, J., Araya, J. F., et al. (2018). Taxonomie gebaseerd op wetenschap is noodzakelijk voor wereldwijde instandhouding. PLoS Biol. 16: e2005075. doi: 10.1371 / journal.pbio.2005075
PubMed Abstract / CrossRef Full Text / Google Scholar
Vieites, D. R., Wollenberg, K. C., Andreonec, F., Köhler, J., Glaw, F., and Vences, M. (2009). Een grote onderschatting van de biodiversiteit van Madagaskar blijkt uit een geïntegreerde amfibie-inventaris. Proc. Natl. Acad. Sci. U. S. A. 106, 8267-8272. doi: 10.1073 / pnas.0810821106
CrossRef Full Text
Vogler, A. P., and de Salle, R. (1994). Diagnosticeren van eenheden van Conservation Management. Nadeel. Biol. 8, 354–363. doi: 10.1046 / j. 1523-1739. 1994.08020354.x
CrossRef Full Text / Google Scholar
Waples, R. S. (1991). Pacifische zalm, Oncorhynchus spp., en de definitie van” soort ” in het kader van de Endangered Species Act. Mar. Vis. Rev. 53, 11-22.
Google Scholar
Waples, R. S. (1995). “Evolutionarily significant units and the conservation of biological diversity under the Endangered Species Act,” in Evolution and The Aquatic Ecosystem: Defining Unique Units in Population Conservation, eds J. L. Nielsen (Bethesda, MD:American Fisheries Society), 8-27.
Google Scholar
Waples, R. S., Gustafson, R. G., Weitkamp, L. A., Myers, J. M., Jjonson, O. W., Busby, P. J., et al. (2001). Kenmerkend voor de diversiteit van zalm uit de Pacific Northwest. J. Fish Biol. 59, 1–41. doi: 10.1111 / j. 1095-8649.2001.tb01376.x
CrossRef Full Text / Google Scholar
Waples, R. S., Nammack, M., Cochrane, J. F., and Hutchings, J. A. (2013). A tale of two acts: endangered species listing practices in Canada en de Verenigde Staten. BioScience 63, 723-734. doi: 10.1093 / bioscience / 63.9.723
CrossRef Full Text / Google Scholar
Waples, R. S., Punt, A. E., and Cope, J. M. (2008). Integratie van genetische gegevens in het beheer van mariene hulpbronnen: hoe kunnen we dat beter doen? Fish 9, 423-449. doi: 10.1111 / j. 1467-2979. 2008. 00303.x
CrossRef Full Text / Google Scholar
Wege, J. A., Thiele, K. R., Shepherd, K. A., Butcher, R., Macfarlane, T. D., and Coates, D. J. (2015). Strategische taxonomie in een biodiverse landschap: een nieuwe benadering van het maximaliseren van instandhoudingsresultaten voor zeldzame en slecht bekende flora. Biodivers. Conserv. 24, 17–32. doi: 10.1007 / s10531-014-0785-4
CrossRef Full Text / Google Scholar
Whitlock, R. (2014). Relaties tussen adaptieve en neutrale genetische diversiteit en ecologische structuur en functioneren: een meta-analyse. J. Ecol. 102, 857–872. doi: 10.1111 / 1365-2745. 12240
PubMed Abstract / CrossRef Full Text / Google Scholar
Wiley, E. O. (1978). Het evolutionaire soortenconcept heroverwogen. Syst. Biol. 27, 17–26. doi: 10.2307 / 2412809
CrossRef Full Text / Google Scholar
Wilson, E. O. (2017). Biodiversiteitsonderzoek vraagt om meer inzet. Nat. Ecol. Evol. 1, 1590–1591. doi: 10.1038 / s41559-017-0360-y
PubMed Abstract / CrossRef Full Text / Google Scholar
Yang, Z. H., and Rannala, B. (2014). Ongeleide afbakening van soorten met behulp van DNA-sequentiegegevens van meerdere loci. Mol. Biol. Evol. 31, 3125–3135. doi: 10.1093 / molbev / msu279
PubMed Abstract / CrossRef Full Text / Google Scholar
Yeates, D. K., Seago, A., Nelson, L., Cameron, S. L., Joseph, L., and Trueman, W. H. (2011). Integratieve taxonomie, of iteratieve taxonomie? Sys. Ent. 36, 209–217. doi: 10.1111/j.1365-3113.2010.00558.x
CrossRef Full Text | Google Scholar