Spierplasticiteit
skeletspieren zijn het grootste weefsel in het lichaam. Het is hoogst plastic en kan dramatische niet-pathologische schommelingen in zowel massa als sterkte in reactie op een groot aantal omgevingsinvloeden ondergaan. Na weerstand oefening of blootstelling aan anabole steroïden, spieren ondergaan hypertrofie en verhoging van de massa, dwarsdoorsnede gebied, en kracht (Aagaard, 2004; Schiaffino et al., 2013). Deze zelfde parameters kunnen neerwaarts afnemen wanneer spieren atrofie in reactie op een van de waaier van beledigingen initiëren die omvatten: honger, denervatie, immobilisatie, sepsis, kanker cachexie, hartfalen, diabetes, nierfalen, chronische obstructieve longaandoening en verworven immunodeficiëntie syndroom (Schiaffino et al., 2013).
de gevolgen van atrofie hebben duidelijke implicaties voor de gezondheid. Spierzwakte is een belangrijke bijdrage aan zowel mortaliteit als morbiditeit en wordt geassocieerd met het verhoogde risico op alle doodsoorzaken (Metter et al., 2002). In feite kan het verminderen van spieratrofie bij kanker cachexie het leven aanzienlijk verlengen (Zhou et al., 2010). Ook, lijden vele oudere individuen aan sarcopenia, een langdurige spier die wanorde verspillen die typisch na de leeftijd van 50 begint en een verlies van ongeveer 1% van spiermassa per jaar impliceert (Woo, 2017). Dit betekent dat door de leeftijd van 80, sarcopenic individuen ongeveer 40% van hun spiermassa, een belangrijke factor in Vallen, broosheid, en verpleeghuis toelatingen hebben verloren. Bijgevolg is het begrijpen van de mechanismen en potentiële therapeutische reacties op atrofie van breed fundamenteel en klinisch belang (Ali and Garcia, 2014; Cohen et al., 2015; Ziaaldini et al., 2017).
Myonucleaire Domeinhypothese
skeletspieren zijn vrij uniek omdat de rijpe cellen syncytieel zijn en honderden kernen kunnen bevatten. Dit is een noodzakelijke aanpassing gezien het feit dat individuele spiervezels enorm kunnen zijn, het bereiken van lengtes tot ~ 600 mm (23 in) (bijvoorbeeld sartorius spier; Yang et al., 1998) en volumes die meer dan 100.000 keer groter kunnen zijn dan een typische mononucleaire cel (Bruusgaard et al., 2003). De bijdragen van veelvoudige kernen worden vereist om grote hoeveelheden mRNA te produceren die nodig zijn om de wonderbaarlijke eiwitsynthese te leiden die wordt vereist om het contractiele apparaat te produceren en te handhaven (Nevalainen et al., 2013).
zowel de plastische aard van spieren als de syncytiële organisatie ervan hebben aanleiding gegeven tot een controverse die pas onlangs lijkt te zijn opgelost—de “myonuclear domain hypothesis” (Qaisar and Larsson, 2014; Gundersen, 2016; Schwartz et al., 2016). Deze theorie heeft zijn oorsprong in het concept van “Wirkungssphäre” of “invloedssfeer” voorgesteld door Strassburger (1893), waarin hij betoogde dat een kern alleen een discreet volume van cytoplasma kan ondersteunen, waardoor de bovengrens van de celgrootte wordt bepaald. Dit principe werd verder uitgewerkt door Gregory (2001) die betoogde dat cellulaire deoxyribonucleïnezuurgehalte en volume nauw zijn gekoppeld. De syncytial aard van skeletachtige spier helpt deze cellen overwinnen deze beperking. De myonucleaire domeinhypothese dicteert dat om de juiste kern-aan-cytoplasmic verhouding te handhaven, nieuwe kernen tijdens hypertrofie worden toegevoegd en met atrofie verloren.er zijn aanzienlijke gegevens die aantonen dat het aantal kernreactoren toeneemt met spierhypertrofie (Moss, 1968; Cabric and James, 1983; Egner et al., 2016), hoewel enige controverse blijft (Murach et al., 2018). Dit proces is een gebied van intensief onderzoek geweest en enkele van de onderliggende moleculaire mechanismen die de verwerving van nieuwe kernen tijdens hypertrofie regelen zijn onlangs gedefinieerd (Guerci et al., 2012; Bentzinger et al., 2014; Ross et al., 2018). Deze supernumeraire kernen worden verworven wanneer lineage-beperkte stamcellen, typisch satellietcellen, fuseren met spiervezels en dragen hun kernen (Brack en Rando, 2012; Bachman et al., 2018). Satellietcellen bevinden zich onder het keldermembraan en aan de sarcolemma (Katz, 1961; Mauro, 1961). Ze blijven rustig tot gestimuleerd door ofwel anabole steroïden zoals testosteron of door focale verwonding na weerstand oefening, op welk punt zij opnieuw de celcyclus ingaan en verspreiden (Joubert and Tobin, 1995; Abreu et al., 2017). Sommige dochtercellen fuseren met de spiervezel en vergemakkelijken zowel reparatie als groei, terwijl anderen de satellietpool arresteren en reconstrueren (Dumont et al., 2015; Goh en Millay, 2017). De resterende overtollige cellen ondergaan apoptose (Schwartz, 2008).
het controversiële aspect van de myonucleaire domeinhypothese is de stelling dat myonuclei verloren gaan tijdens atrofie. Er zijn vele interventies die atrofie in dierlijke modellen veroorzaken, met inbegrip van: immobilisatie, denervatie, en sepsis (Fitts et al., 1986; Minnaard et al., 2005; O ‘ Leary et al., 2012). In elk geval is er een netto verlies van zowel spier dwarsdoorsnede gebied (de primaire assay) en het verschijnen van apoptotische cellen in het weefsel (McCall et al., 1998; Smith et al., 2000; Strasser et al., 2000; Alway et al., 2003; McClung et al., 2007; Andrianjafiniony et al., 2010; Guo et al., 2012; Palumbo et al., 2012; Barnes et al., 2015; Cheema et al., 2015; Li et al., 2016; Kletzien et al., 2018). In deze studies, wordt apoptose gemeten in een verscheidenheid van methodes, met inbegrip van caspaseactivering, mitochondrial endog versie, of de fragmentatie van DNA . Deze studies verstrekken dwingende gegevens die apoptosis dramatisch tijdens de vroege fase van atrofie toeneemt. Bijvoorbeeld, in een recente uitgebreide studie (Guo et al., 2012), Guo et al. onderworpen muizen aan 14 dagen van achterpoten suspensie, een behandeling die resulteerde in een ~69% vermindering van spier nat gewicht en een ~43,8% vermindering van dwarsdoorsnede gebied ten opzichte van de ongemanipuleerde contralaterale spier. De auteurs gekwantificeerd ook een aantal apoptose markers, met inbegrip van TUNEL kleuring, caspase-3 splitsing / activering, en de splitsing van poly(adenosine difosfaat ribose) polymerase (PARP), een eiwit betrokken bij DNA-reparatie. Elk van deze apoptose-maatregelen nam significant toe na de achterpootsuspensie. Terwijl de primaire focus van dit artikel was om de positieve impact van elektrische stimulatie op het beperken van atrofie na een belediging te evalueren, zij en vele andere onderzoekers interpreteren dit soort gegevens als het verstrekken van sterke ondersteuning voor de myonucleaire domein hypothese.
vanuit een celbiologisch perspectief vormt het vermoedelijke verlies van kernen binnen een syncytieel weefsel zoals skeletspieren een groot praktisch probleem. Hoe kan een individuele kern zo gecompromitteerd worden dat zijn genoom snel condenseert en fragmenteert terwijl zijn buren blijven bestaan en de levensvatbaarheid van de spiervezel helpen behouden? Aangezien apoptose typisch wordt gemedieerd door de activering van de klasse van cysteine proteasen bekend als caspasen, is het niet duidelijk welk mechanisme zou kunnen dienen om de activiteit van een diffuseerbaar protease binnen een gemeenschappelijk cytoplasma te beperken.
deze vraag is indirect behandeld in een ander syncytieel celtype, het menselijke syncytiotrofoblast, een weefsel dat de placenta omringt en ongeveer 5 × 1010 kernen bevat (Mayhew et al., 1999). Wanneer apoptose wordt geïnduceerd in het syncytiotrofoblast, verspreidt het zich als een golf met een snelheid van ongeveer 5 micron per minuut totdat het hele weefsel is betrokken (Longtine et al., 2012). Bijgevolg zijn er geen” bevoorrechte ” gebieden binnen het syncytieel cytoplasma en worden alle kernen uiteindelijk vernietigd.
een van de belangrijkste uitdagingen bij het analyseren van apoptose in skeletspieren is dat het een zeer heterogeen weefsel is, waar ongeveer de helft van zijn kernen zich buiten spiervezels bevinden (Schmalbruch en Hellhammer, 1977). Deze mononucleated cellen omvatten satellietcellen, endothelial cellen, fibroblasten, pericytes ,en macrofagen (Tedesco et al., 2010). Daarom is het erg moeilijk om te bepalen welke kant van het sarcolemma, een kern, zich bevindt, en dus of het een echte myonucleus is.
in Vivo Time-Lapse beeldvorming van gelabelde Muisspiervezels
ondanks het grote aantal papers die apoptose tijdens spieratrofie aantonen, hebben verscheidene auteurs deze resultaten in twijfel getrokken (Wada et al., 2002; Zhong et al., 2005; Aravamudan et al., 2006; Gundersen and Bruusgaard, 2008; Duddy et al., 2011; Qaisar en Larsson, 2014). Bijvoorbeeld, met behulp van geïsoleerde spiervezels in vitro, duddy et al. opgemerkt dat terwijl spiervezel volume daalde in de tijd, reflecterend van een atrofisch proces, het aantal myonuclei leek stabiel te zijn (Duddy et al., 2011).
in een reeks elegante experimenten injecteerde het Gundersen lab individuele extensor digitorum longus (EDL) of soleus spieren spiervezels met kleurstoffen die onafhankelijk de kernen en het cytoplasma labelden in verdoofde muizen (Bruusgaard and Gundersen, 2008; Bruusgaard et al., 2010). Dit stond hen toe om elke kern binnen individuele spiervezels in vivo in tijd te identificeren en dan zijn lot te evalueren. Bijvoorbeeld, EDL spieren werden veroorzaakt tot hypertrofie door de ablatie van hun belangrijkste synergists (Bruusgaard et al., 2010). Tussen dag 6 en 11 nam het aantal myonuclei toe met ongeveer 54% en tussen dag 9 en 14 was er een 35% toename van de dwarsdoorsnede (figuur 1). Deze gegevens zijn consistent met de hypothese dat spieren supernumeraire kernen verwerven voorafgaand aan de belangrijke groei van de vezel tijdens hypertrofie.

figuur 1. Myonuclei worden verworven tijdens hypertrofie maar niet verloren tijdens atrofie bij muis. Micrografen van dezelfde EDL spiervezel na verloop van tijd na de inductie van hypertrofie (bovenste rij) en de daaropvolgende inductie van atrofie (onderste rij). Fluorescently geëtiketteerde oligonucleotides werden gebruikt om de kernen in vivo te visualiseren. De stippellijnen stellen het sarcolemma voor. Schaalbalk = 50 µm. (Aangepast van Bruusgaard et al., 2010. Gebruikt met toestemming van de Proceedings van de National Academy of Sciences.)
zij onderzochten vervolgens het lot van deze uniek identificeerbare myonuclei in dezelfde spiervezels wanneer zij werden geïnduceerd atrofie te ondergaan. Denervatie leidde tot een vermindering van ongeveer 50% van het spiervezelvolume, maar geen verlies van myonuclei (figuur 1; Bruusgaard et al., 2010). Het niet waarnemen van nucleair verlies was niet te wijten aan de atrofische stimulus die werd gebruikt, omdat ze dezelfde resultaten verkregen wanneer de spieren werden geïnduceerd tot atrofie in reactie op tetrodotoxine-geïnduceerde zenuwblokkade, achterpoten suspensie, kanker cachexie, of detraining (Bruusgaard and Gundersen, 2008; Bruusgaard et al., 2010; Winje et al., 2018a). In feite, in de loop van hun studies, ze onderzocht meer dan 200.000 individuele myonuclei in atrofische spieren en waargenomen slechts 4 TUNEL-positieve (apoptotische) kernen, die een verlies van slechts ~0,002% van de kernen vertegenwoordigt (Bruusgaard et al., 2012). Deze gegevens ondersteunen sterk de hypothese dat skeletspieratrofie niet gepaard gaat met myonucleaire dood.
Insectenspier als Model
zo krachtig als deze studies zijn, zijn er enkele beperkingen inherent aan het gebruik van zoogdiermodellen. Ten eerste, Vertrouwen zij steevast op experimentele interventies zoals denervatie om spieratrofie te veroorzaken. Ten tweede, wordt de contralaterale spier typisch gebruikt als interne controle hoewel het ook aan sommige van de zelfde milieusignalen, zoals spanningshormonen wordt blootgesteld (Bonaldo en Sandri, 2013). Ten slotte kunnen de skeletachtige spieren geprogrammeerde celdood (PCD) tijdens ontwikkeling ook ondergaan, en deze modellen richten zelden de rol van apoptosis in deze cellen aan.
een alternatief model dat aan geen van deze beperkingen lijdt en het mogelijk maakt spierkernen te bestuderen tijdens zowel atrofie als natuurlijk voorkomende PCD, is de intersegmentale spier (ISM) van de Tobacco hawkmoth Manduca sexta. De ISMs zijn samengesteld uit platen van gigantische spiervezels, waarbij elke cel is ongeveer 5 mm lang en tot 1 mm in diameter (figuur 2A). De ismen hechten zich aan de segmentale grenzen binnen het achterlijf en genereren zowel het kruipende gedrag van de larven als het eclosiegedrag van de volwassen mot wanneer deze aan het einde van de metamorfose uit de poppenhuid ontsnapt.
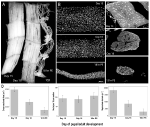
Figuur 2. Retentie van myonuclei tijdens zowel atrofie als dood van de intersegmentale spieren (ISMs) van de mot Manduca sexta. (A) ismen uit drie ontwikkelingsstadia: homeostatisch (dag 15 van de ontwikkeling van de pop-adult; links); atrofisch (dag 18; Midden); en sterven (18 uur na eclosie; rechts). Schaalbalk is gelijk aan ~1 mm (aangepast van Schwartz et al., 2016). (B) ISMs van deze zelfde ontwikkelingsstadia werden gezuiverd en bevlekt met de nucleaire kleurstof DAPI en gevisualiseerd via confocale microscopie (aangepast van Schwartz et al., 2016). (C) ISM-vezelsecties (10 µm) werden gekleurd met de nucleaire kleurstof DAPI. Let op het dramatische verlies van spiereiwit (lichtgrijs gebied) tijdens atrofie en dood, maar het behoud van kernen in alle stadia (aangepast van Schwartz et al., 2016). (D) kwantificering van ISM-vezelvolume (links), kerngetal (Midden) en myonucleaire domeingrootte (rechts) tijdens homeostase, atrofie en dood. (Gemiddelde ± standaardfout.) (Aangepast van Schwartz et al., 2016).
Op dag 15 van de normale 18 dagen van ontwikkeling van pop-volwassenen, starten de ISMs een hormonaal getriggerd atrofieprogramma dat resulteert in een verlies van 40% van de massa bij eclosie 3 dagen later (figuur 2A; Schwartz and Truman, 1983). Dit dramatische verlies van spiermassa is gelijk aan dat gezien in 80-jarige mensen met sarcopenia, maar vindt plaats meer dan 3 dagen in plaats van 30 jaar. Terwijl de ISMs massa verliezen, behouden zij normale fysiologische eigenschappen zoals rustpotentieel en kracht / dwarsdoorsnede gebied (Schwartz and Ruff, 2002). De ISMs worden gebruikt om het eclosiegedrag aan het eind van dag 18 te produceren en dan PCD in werking te stellen, die beide door een peptide hormoon worden teweeggebracht (Schwartz en Truman, 1982, 1983). .in tegenstelling tot zoogdierspieren bevatten de ISMs geen capillairen, satellietcellen, endotheelcellen of pericyten, wat betekent dat vrijwel alle kernen in het weefsel zich in de spiervezels zelf bevinden (Beaulaton en Lockshin, 1977). We gebruikten twee onafhankelijke methoden om het nucleaire lot tijdens atrofie en dood in dit model te controleren (Schwartz et al., 2016). De eerste was een standaard anatomische benadering. Wanneer de ISMs atrofie en dood ondergaan, zijn er dramatische verminderingen in zowel het volume als de dwarsdoorsnede gebied van de vezels, maar grofweg, kernaantal onveranderd was (figuur 2B). Met behulp van gesegmenteerde weefsels voor kwantitatieve studies, vonden we dat er een ~49% vermindering van vezel dwarsdoorsnede gebied tijdens atrofie en een verdere 30% daling tijdens de vroege stadia van de dood (figuren 2C,d; Schwartz et al., 2016). Het aantal kernen veranderde echter niet tijdens dezelfde periode, wat betekende dat er functioneel een vermindering van 84% was in het myonucleaire domein.
de tweede methode die we gebruikten was het meten van het DNA-gehalte van individuele spiervezels van dieren op dag 13, wanneer de spieren homeostatisch zijn, tot 18 uur na eclosie, wanneer de spieren sterk gedegenereerd zijn (Schwartz et al., 2016). We kwantificeerden het DNA-gehalte van 420 individuele cellen en vonden dat er geen significant verlies was tijdens de ontwikkeling. Deze biochemische analyses stemmen Goed met de anatomische gegevens overeen en stellen voor dat de relatieve concentratie van zowel kernen als genomic DNA functioneel toenam aangezien de spieren atrofieerden en stierven. Bijgevolg suggereren gegevens van zowel muizen als motten dat spierkernen geen apoptose ondergaan tijdens atrofie (of PCD) en leiden tot de conclusie dat de myonucleaire domeinhypothese moet worden afgewezen.
conclusies
deze waarnemingen hebben een aantal implicaties voor zowel het begrijpen van de fundamentele biologie van spieren als voor het ontwikkelen van mogelijke therapeutische interventies. Terwijl er substantiële gegevens rapportage van de aanwezigheid van apoptotische kernen binnen het weefsel na atrofische beledigingen, recente gegevens suggereren dat deze niet waar zijn myonuclei, maar eerder, veroordeeld mononucleaire cellen die buiten de spiervezel wonen. De belangrijkste reden voor deze discrepantie is dat de traditionele instrumenten die worden gebruikt voor het opsporen van apoptose niet de resolutie die nodig is om adequaat te bepalen welke kant van de sarcolemma een stervende cel zich bevindt. Methoden die specifiek myonuclei labelen, hetzij via in vivo injecties van fluorescente kleurstoffen (Bruusgaard en Gundersen, 2008) of via genetische manipulaties om markerproteïnen tot expressie te brengen (Duddy et al., 2011), onthullen weinig of geen apoptotische kernen binnen de geëtiketteerde vezels. De recente demonstratie dat antilichamen gericht tegen eiwit Pericentriolair materiaal 1 (pcm1) label alleen authentieke myonuclei biedt een veelbelovende tool die kan worden toegepast op de traditionele histologische materialen die routinematig worden geanalyseerd in het veld (Winje et al., 2018b).
Het is niet verwonderlijk dat de skeletspieren apoptose gebruiken als mechanisme voor het elimineren van myonuclei. Het lijkt erop dat Rijpe skeletspieren grotendeels uitgesloten zijn van het initiëren van apoptose omdat het de expressie van krachtige overlevingseiwitten zoals X-gebonden remmer van apoptose-eiwit (Xiap) upreguleert (Smith et al., 2009) en apoptose repressor met caspase recruitment domain (ARC) (Xiao et al., 2011), terwijl tegelijkertijd lage niveaus van apoptosis mediatoren zoals apoptotische protease activerende factor 1 (Apaf-1) (Burgess et al., 1999). Dit is zinvol teleologisch gezien spieren zijn onderworpen aan extreme verstoringen, met inbegrip van verstoring van de sarcolemma na intense oefening, dus het zou catastrofaal zijn als ze celdood in plaats van geïnitieerde weefselreparatie en mogelijk hypertrofie. Net als andere terminaal gedifferentieerde cellen, hebben spieren de neiging om niet-apoptotische mechanismen in te zetten, het meest in het bijzonder autophagic PCD (ook bekend als type II degeneratie) (Clarke, 1990; Schwartz et al., 1993; Kole et al., 2013; Ginet et al., 2014) wanneer ze sterven tijdens de ontwikkeling.
zelfs als er een mechanisme was waardoor spieren selectief individuele kernen konden richten, is het niet duidelijk dat dit nuttig zou zijn. In feite, verleent het behoud van” surplus ” kernen tijdens atrofie een duidelijk voordeel voor het individu aangezien skeletspieren vaak cycli van atrofie en hypertrofie in reactie op milieuvoorwaarden zoals voedselbeschikbaarheid ondergaan. Het vermogen om snel te herstellen door gebruik te maken van reeds bestaande myonuclei kan een belangrijke rol spelen bij de aanpassing (Jackson et al., 2012) en helpen het fenomeen van “spiergeheugen” (Staron et al., 1991; Gundersen, 2016). Het is goed gedocumenteerd op het gebied van de oefeningsfysiologie dat het veel gemakkelijker is om een bepaald niveau van spiergeschiktheid door oefening te verwerven dan het was om het te bereiken de eerste plaats, zelfs als er een lange tussenliggende periode van detraining is geweest. Met andere woorden, de uitdrukking “use it or lose it” is misschien nauwkeuriger geformuleerd als “use it or lose it, until you work at it again.”Dit is direct aangetoond door een ander experiment van het Gundersen lab dat aantoonde dat zodra een spier nieuwe kernen heeft verworven, het deze vasthoudt lang nadat de hypertrofische stimulus is verwijderd. Zij induceerden spierhypertrofie in vrouwtjesmuizen door hen gedurende 2 weken met testosteron te behandelen en vervolgens de spieren 3 weken na Steroid terugtrekking (Egner et al., 2013). Spiervolume was teruggekeerd naar baseline, maar de nieuw verworven kernen bleven zelfs 3 maanden later. Wanneer de spieren werden onderworpen aan overbelasting om hypertrofie te herstarten, ondergingen de steroïde-behandelde degenen snel een 36% verhoging van het vezelvolume terwijl de controle spieren slechts met 6% groeide. Deze gegevens suggereren dat de” overtollige ” kernen snel zouden kunnen worden gemobiliseerd om herscholing te vergemakkelijken.
deze waarnemingen hebben mogelijke implicaties voor het volksgezondheidsbeleid. Het is aangetoond dat spiergroei, fysiologische functie en regeneratieve capaciteit allemaal afnemen met de leeftijd, grotendeels als gevolg van verminderde satellietcelproliferatie (Blau et al., 2015). Bijgevolg, zou de oefening tijdens adolescentie, wanneer de spiergroei door hormonen, voeding en een robuuste satellietpool wordt verbeterd, functioneel kunnen dienen om individuen aan “bank” myonuclei toe te staan die op later in het leven zou kunnen worden getrokken om de gevolgen van het verouderen te vertragen en misschien sarcopenia te voorkomen.
bovendien hebben deze gegevens implicaties op het gebied van competitieve sporten. Het gebruik van anabole steroïden is een krachtige stimulans voor spierhypertrofie en de toevoeging van nieuwe myonuclei (Egner et al., 2013). Aangezien deze kernen lang na de einden van het steroidgebruik blijven bestaan, ontlenen de atleten waarschijnlijk de voordelen van illegaal druggebruik zonder het risico van opsporing.
samenvattend kan worden gesteld dat hoewel de toevoeging van nieuwe kernen met spiergroei grotendeels wordt aanvaard, het apoptotisch verlies van kernen met atrofie niet kan worden ondersteund, hetgeen erop wijst dat een strikte interpretatie van de myonucleaire domeinhypothese niet kan worden ondersteund. In plaats daarvan, lijkt het erop dat zodra verworven, myonuclei zelfs wanneer een spier atrofisch wordt of celdood in werking stelt volhardt.
Auteursbijdragen
de auteur bevestigt de enige bijdrager van dit werk te zijn en heeft het goedgekeurd voor publicatie.
financiering
het werk voor deze publicatie werd ondersteund door fondsen van de Eugene M. en Ronnie Isenberg hoogleraarschap Endowment. Er werd geen bedrijfsondersteuning aan de auteur verstrekt en er worden geen commerciële producten vermeld of onderschreven.
belangenconflict verklaring
de auteur verklaart dat het onderzoek werd uitgevoerd zonder commerciële of financiële relaties die als een potentieel belangenconflict kunnen worden opgevat.Dankbetuigingen de auteur wil mevrouw Christine Brown bedanken voor de hulp bij de voorbereiding van dit manuscript en de Eugene M. en Ronnie Isenberg Lectorship Endowment voor ondersteuning.
Aagaard, P. (2004). Spieren “sterker” maken: lichaamsbeweging, voeding, drugs. J. Musculoskelet. Neuronaal. Interactie. 4, 165–174.
PubMed Abstract / Google Scholar
Abreu, P., Mendes, S. V., Ceccatto, V. M., and Hirabara, S. M. (2017). Satellietcelactivering geïnduceerd door aërobe spieraanpassing als reactie op uithoudingsvermogen bij mensen en knaagdieren. Life Sci. 170, 33–40. doi: 10.1016 / j. lfs.2016.11.016
PubMed Abstract / CrossRef Full Text / Google Scholar
Ali, S., and Garcia, J. M. (2014). Sarcopenie, cachexie en veroudering: diagnose, mechanismen en therapeutische opties—een mini-review. Gerontology 60, 294-305. doi: 10.1159 / 000356760
PubMed Abstract / CrossRef Full Text / Google Scholar
altijd, S. E., Martyn, J. K., Ouyang, J., Chaudhrai, A., and Murlasits, Z. S. (2003). Id2 expression during apoptosis and satellite cell activation in unloaded and loaded quail skeletal muscles. Am. J. Physiol. Regul. Integr. Comp. Physiol. 284, R540–R549. doi: 10.1152/ajpregu.00550.2002
PubMed Abstract | CrossRef Full Text | Google Scholar
Andrianjafiniony, T., Dupre-Aucouturier, S., Letexier, D., Couchoux, H., and Desplanches, D. (2010). Oxidative stress, apoptosis, and proteolysis in skeletal muscle repair after unloading. Am. J. Phys. Cell Physiol. 299, C307–C315. doi: 10.1152/ajpcell.00069.2010
PubMed Abstract / CrossRef Full Text / Google Scholar
Aravamudan, B., Mantilla, C. B., Zhan, W. Z., and Sieck, G. C. (2006). Denervatie effecten op myonucleaire domeingrootte van rat diafragma vezels. J. Appl. Fysiol. 100, 1617–1622. doi: 10.1152 / japplphysiol.01277.2005
CrossRef Full Text / Google Scholar
Bachman, J. F., Klose, A., Liu, W., Paris, N. D., Blanc, R. S., Schmalz, M., et al. (2018). Prepuberale skeletspiergroei vereist pax 7-expressie satellietcel-afgeleide myonucleaire bijdrage. Ontwikkeling 145: 167197.
Google Scholar
Barnes, B. T., Confides, A. L., Rich, M. M., and Dupont-Versteegden, E. E. (2015). De verschillende spierapoptotische wegen worden geactiveerd in spieren met verschillende vezel types in een rattenmodel van kritieke ziekte myopathie. J. Muscle Res. Cell Motil. 36, 243–253. doi: 10.1007/s10974-015-9410-8
PubMed Abstract / CrossRef Full Text/Google Scholar
Beaulaton, J., and Lockshin, R. A. (1977). Ultrastructurele studie van de normale degeneratie van de intersegmentale spieren van Anthereae polyphemus en Manduca sexta (insecta, lepidoptera) met bijzondere verwijzing naar cellulaire autophagie. J. Morphol. 154, 39–57. doi: 10.1002 / jmor.1051540104
PubMed Abstract / CrossRef Full Text/Google Scholar
Bentzinger, C. F., von Maltzahn, J., Dumont, N. A., Stark, D. A., Wang, Y. X., Nhan, K., et al. (2014). Wnt 7a stimuleert myogene stamcelmotiliteit en engraftment wat resulteert in verbeterde spierkracht. J. Cell Biol. 205, 97–111. doi: 10.1083 / jcb.201310035
PubMed Abstract / CrossRef Full Text / Google Scholar
Blau, H. M., Cosgrove, B. D., and Ho, A. T. (2015). De centrale rol van spierstamcellen in regeneratieve mislukking met het verouderen. Nat. Med. 21, 854–862. doi: 10.1038 / nm.3918
PubMed Abstract / CrossRef Full Text/Google Scholar
Bonaldo, P., and Sandri, M. (2013). Cellulaire en moleculaire mechanismen van spieratrofie. Dis. Model. Mech. 6, 25–39. doi: 10.1242/dmm.010389
CrossRef Full Text / Google Scholar
Brack, A. S., and Rando, T. A. (2012). Weefsel-specifieke stamcellen: lessen uit de skeletspier satellietcel. Cel Stamcel 10, 504-514. doi: 10.1016 / j.stam.2012.04.001
PubMed Abstract / CrossRef Full Text / Google Scholar
Bruusgaard, J. C., Egner, I. M., Larsen, T. K., Dupre-Aucouturier, S., Desplanches, D., and Gundersen, K. (2012). Geen verandering in het myonucleaire aantal tijdens het lossen en herladen van spieren. J. Appl. Fysiol. 113, 290–296. doi: 10.1152 / japplphysiol.00436.2012
PubMed Abstract / CrossRef Full Text / Google Scholar
Bruusgaard, J. C., and Gundersen, K. (2008). In vivo time-lapse microscopie onthult geen verlies van muriene myonuclei tijdens weken van spieratrofie. J. Clin. Investeren. 118, 1450–1457. doi: 10.1172 / JCI34022
PubMed Abstract / CrossRef Full Text / Google Scholar
Bruusgaard, J. C., Johansen, I. B., Egner, I. M., Rana, Z. A., and Gundersen, K. (2010). Myonuclei verkregen door overbelasting oefening gaat vooraf aan hypertrofie en zijn niet verloren bij detraining. Proc. Natl. Acad. Sci. U. S. A. 107, 15111-15116. doi: 10.1073 / pnas.0913935107
CrossRef Full Text / Google Scholar
Bruusgaard, J. C., Liestol, K., Ekmark, M., Kollstad, K., and Gundersen, K. (2003). Aantal en ruimtelijke verdeling van kernen in de spiervezels van normale muizen onderzocht in vivo. J. Physiol. 551, 467–478. doi: 10.1113 / jphysiol.2003.045328
PubMed Abstract / CrossRef Full Text / Google Scholar
Burgess, D. H., Svensson, M., Dandrea, T., Gronlund, K., Hammarquist, F., Orrenius, S., et al. (1999). Cytosolen van menselijke skeletspieren zijn refractair voor cytochroom C-afhankelijke activering van type-II caspasen en missen APAF-1. Celdood Verschilt. 6, 256–261. doi: 10.1038 / sj.cdd.4400489
PubMed Abstract / CrossRef Full Text/Google Scholar
Cabric, M., and James, N. T. (1983). Morfometrische analyses op de spieren van training getrainde en ongetrainde honden. Is. J. Anat. 166, 359–368. doi: 10.1002 / aja.1001660309
PubMed Abstract / CrossRef Full Text / Google Scholar
Cheema, N., Herbst, A., McKenzie, D., and Aiken, J. M. (2015). Apoptose en necrose bemiddelen het verlies van skeletspieren in door leeftijd geïnduceerde mitochondriale enzymatische afwijkingen. Aging Cell 14, 1085-1093. doi: 10.1111 / acel.12399
PubMed Abstract / CrossRef Full Text/Google Scholar
Clarke, P. G. (1990). Ontwikkeling celdood: morfologische diversiteit en meerdere mechanismen. Anat. Embryol. 181, 195–213.
Google Scholar
Cohen, S., Nathan, J. A., and Goldberg, A. L. (2015). Spier verspillen bij ziekte: moleculaire mechanismen en veelbelovende therapieën. Nat. Rev. Drug Discov. 14, 58–74. doi: 10.1038 / nrd4467
PubMed Abstract / CrossRef Full Text / Google Scholar
Duddy, W. J., Cohen, T., Duguez, S., and Partridge, T. A. (2011). De geïsoleerde spiervezel als model van onbruik atrofie: karakterisering gebruikend PhAct, een methode om f-actin te kwantificeren. Exp. Cel Res. 317, 1979-1993. doi: 10.1016 / j.yexcr.2011.05.013
PubMed Abstract / CrossRef Full Text / Google Scholar
Dumont, N. A., Wang, Y. X., and Rudnicki, M. A. (2015). Intrinsieke en extrinsieke mechanismen die satelliet celfunctie regelen. Ontwikkeling 142, 1572-1581. doi: 10.1242 / dev.114223
PubMed Abstract / CrossRef Full Text / Google Scholar
Egner, I. M., Bruusgaard, J. C., Eftestol, E., and Gundersen, K. (2013). Een cellulair geheugen mechanisme helpt overbelasting hypertrofie in spier lang na een episodische blootstelling aan anabole steroïden. J. Physiol. 591, 6221–6230. doi: 10.1113 / jphysiol.2013.264457
PubMed Abstract / CrossRef Full Text / Google Scholar
Egner, I. M., Bruusgaard, J. C., and Gundersen, K. (2016). Satellietceldepletie voorkomt vezelhypertrofie in skeletspieren. Ontwikkeling 143, 2898-2906. doi: 10.1242 / dev.134411
PubMed Abstract / CrossRef Full Text/Google Scholar
Fitts, R. H., Metzger, J. M., Riley, D. A., and Unsworth, B. R. (1986). Modellen van onbruik: een vergelijking van achterpootopschorting en immobilisatie. J. Appl. Fysiol. 60, 1946–1953.
Google Scholar
Ginet, V., Pittet, M. P., Rummel, C., Osterheld, M. C., Meuli, R., Clarke, P. G., et al. (2014). Stervende neuronen in thalamus van gestikte term pasgeborenen en ratten zijn autofagisch. Anne. Neurol. 76, 695–711. doi: 10.1002 / ana.24257
PubMed Abstract / CrossRef Full Text / Google Scholar
Goh, Q., and Millay, D. P. (2017). Vereiste van myomaker-gemedieerde stamcelfusie voor skeletspierhypertrofie. elife 6. doi: 10.7554 / eLife.20007
PubMed Abstract / CrossRef Full Text/Google Scholar
Gregory, T. R. (2001). Toeval, coevolutie of oorzakelijk verband? DNA-inhoud, celgrootte en de C-waarde enigma. Biol. Rev. Camb. Philos. Soc. 76, 65–101. doi: 10.1017 / s1464793100005595
PubMed Abstract / CrossRef Full Text / Google Scholar
Guerci, A., Lahoute, C., Hebrard, S., Collard, L., Graindorge, D., Favier, M., et al. (2012). Srf-afhankelijke paracrine signalen geproduceerd door myofibers controle satelliet cel-gemedieerde skeletachtige spier hypertrofie. Cel Metab. 15, 25–37. doi: 10.1016 / j. cmet.2011.12.001
PubMed Abstract / CrossRef Full Text / Google Scholar
Gundersen, K. (2016). Spiergeheugen en een nieuw cellulair model voor spieratrofie en hypertrofie. J. Exp. Biol. 219, 235–242. doi: 10.1242/jeb.124495
PubMed Abstract / CrossRef Full Text / Google Scholar
Gundersen, K., and Bruusgaard, J. C. (2008). Nucleaire domeinen tijdens spieratrofie: kernen verloren of paradigma verloren? J. Physiol. 586, 2675–2681. doi: 10.1113 / jphysiol.2008.154369
PubMed Abstract / CrossRef Full Text / Google Scholar
Guo, B. S., Cheung, K. K., Yeung, S. S., Zhang, B. T., and Yeung, E. W. (2012). Elektrische stimulatie beïnvloedt satellietcelproliferatie en apoptose in lossingsgeïnduceerde spieratrofie bij muizen. PLoS ONE 7: e30348. doi: 10.1371 / journal.pone.0053523
PubMed Abstract / CrossRef Full Text/Google Scholar
Jackson, J. R., Mula, J., Kirby, T. J., Fry, C. S., Lee, J. D., Ubele, M. F., et al. (2012). Depletie van satellietcellen remt de hergroei van volwassen skeletspieren na lossing-geïnduceerde atrofie niet. Is. J. Phys. Celfysiol. 303, C854-C861. doi: 10.1152/ajpcell.00207.2012
PubMed Abstract / CrossRef Full Text / Google Scholar
Joubert, Y., and Tobin, C. (1995). De testosteronbehandeling resulteert in rustige satellietcellen die in celcyclus in rat levator ani spier worden geactiveerd en gerekruteerd. Dev. Biol. 169, 286–294. doi: 10.1006 / dbio.1995.1144
PubMed Abstract / CrossRef Full Text/Google Scholar
Katz, B. (1961). De terminaties van de afferente zenuwvezel in de spierspil van de kikker. Philos. Transvetzuren. R. Soc. Lond. B Biol. Sci. 243, 221–240.
PubMed Abstract / Google Scholar
Kletzien, H., Hare, A. J., Leverson, G., and Connor, N. P. (2018). Leeftijdsgebonden effect van celdood op de vezelmorfologie en het aantal in de tongspier. Spierzenuw 57, E29-E37. doi: 10.1002/mus.25671
PubMed Abstract / CrossRef Full Text / Google Scholar
Kole, A. J., Annis, R. P., and Deshmukh, M. (2013). Volwassen neuronen: uitgerust om te overleven. Celdood Dis. 4: e689. doi: 10.1038 / cddis.2013.220
PubMed Abstract / CrossRef Full Text / Google Scholar
Li, T. S., Shi, H., Wang, L., and Yan, C. (2016). Effect van mesenchymale stamcellen van het beenmerg op de proliferatie van satellietcellen en apoptose in immobilisatie-geïnduceerde spieratrofie bij ratten. Med. Sci. Monit. 22, 4651–4660. doi: 10.12659/MSM.898137
PubMed Abstract / CrossRef Full Text / Google Scholar
Lockshin, R. A., and Williams, C. M. (1965). Geprogrammeerde celdood – I. cytologie van degeneratie in de intersegmentale spieren van de pernyi-zijde. J. Insectenfysiol. 11, 123–133. doi: 10.1016/0022-1910(65)90099-5
PubMed Abstract / CrossRef Full Text / Google Scholar
Longtine, M. S., Barton, A., Chen, B., and Nelson, D. M. (2012). Live-cell imaging toont apoptose initieert lokaal en verspreidt zich als een golf door syncytiotrophoblasten in primaire culturen van menselijke placenta villous trofoblasten. Placenta 33, 971-976. doi: 10.1016 / j.placenta.2012.09.013
PubMed Abstract / CrossRef Full Text / Google Scholar
Mauro, A. (1961). Satellietcel van skeletspiervezels. J. Biophys. Biochem. Cytol. 9, 493–495.
Google Scholar
Mayhew, T. M., Leach, L., McGee, R., Ismail, W. W., Myklebust, R., and Lammiman, M. J. (1999). Proliferatie, differentiatie en apoptose in villous trophoblast bij 13-41 weken zwangerschap (inclusief waarnemingen op annulate lamellen en nucleaire poriecomplexen). Placenta 20, 407-422. doi: 10.1053 / plac.1999.0399.
PubMed Abstract / CrossRef Full Text/Google Scholar
McCall, G. E., Allen, D. L., Linderman, J. K., Grindeland, R. E., Roy, R. R., Mukku, V. R., et al. (1998). Behoud van myonucleaire domeingrootte in rat soleus na overbelasting en groeihormoon / IGF – I behandeling. J. Appl. Fysiol. 84, 1407–1412.
Google Scholar
McClung, J. M., Kavazis, A. N., DeRuisseau, K. C., Falk, D. J., Deering, M. A., Lee, Y., et al. (2007). Caspase-3 regulering van het myonucleaire domein van het middenrif tijdens mechanische ventilatie-geïnduceerde atrofie. Is. J. Respir. Crit. Care Med. 175, 150–159. doi: 10.1164 / rccm.200601-142OC
PubMed Abstract / CrossRef Full Text / Google Scholar
Metter, E. J., Talbot, L. A., Schrager, M., and Conwit, R. (2002). Skeletspiersterkte als een voorspeller van alle-oorzaak mortaliteit bij gezonde mannen. J. Gerontol. Een Biol. Sci. Med. Sci. 57, B359-B365. doi: 10.1093 / gerona / 57.10.B359
PubMed Abstract / CrossRef Full Text/Google Scholar
Minnaard, R., Drost, M. R., Wagenmakers, A. J., Van Kranenburg, G. P., Kuipers, H., and Hesselink, M. K. (2005). Skeletspieren verspillen en contractiele prestaties bij septische ratten. Spierzenuw 31, 339-348. doi: 10.1002/mus.20268
PubMed Abstract / CrossRef Full Text / Google Scholar
Moss, F. P. (1968). De verhouding tussen de afmetingen van de vezels en het aantal kernen bij de normale groei van de skeletspieren bij het pluimvee. Is. J. Anat. 122, 555–563. doi: 10.1002 / aja.1001220308
PubMed Abstract / CrossRef Full Text / Google Scholar
Murach, K. A., Englund, D. A., Dupont-Versteegden, E. E., McCarthy, J. J., and Peterson, C. A. (2018). Myonuclear domain flexibility daagt rigide veronderstellingen over satellietcel bijdrage aan skeletspier vezel hypertrofie. Voorkant. Fysiol. 9:635. doi: 10.3389 / fphys.2018.00635
CrossRef Full Text / Google Scholar
Nevalainen, M., Kaakinen, M., and Metsikko, K. (2013). Distributie van mRNA transcripten en vertaling activiteit in skeletachtige myofibers. Cell Tissue Res. 353, 539-548. doi: 10.1007/s00441-013-1659-x
PubMed Abstract / CrossRef Full Text / Google Scholar
O ‘ Leary, M. F., Vainshtein, A., Carter, H. N., Zhang, Y., and Hood, D. A. (2012). Denervatie-geïnduceerde mitochondriale dysfunctie en autofagie in skeletspieren van apoptose-deficiënte dieren. Is. J. Phys. Celfysiol. 303, C447–C454. doi: 10.1152/ajpcell.00451.2011
PubMed Abstract | CrossRef Full Text | Google Scholar
Palumbo, C., Rovesta, C., and Ferretti, M. (2012). Striated muscle fiber apoptosis after experimental tendon lesion in a rat model. J. Anat. 221, 358–363. doi: 10.1111/j.1469-7580.2012.01554.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Qaisar, R., and Larsson, L. (2014). What determines myonuclear domain size? Indian J. Physiol. Pharmacol. 58, 1–12.
PubMed Abstract | Google Scholar
Ross, J. A., Levy, Y., Svensson, K., Philp, A., Schenk, S., and Ochala, J. (2018). SIRT1 reguleert het nucleaire aantal en de domeingrootte in skeletspiervezels. J. Cell. Fysiol. 233, 7157–7163. doi: 10.1002 / jcp.26542
PubMed Abstract / CrossRef Full Text / Google Scholar
Schiaffino, S., Dyar, K. A., Ciciliot, S., Blaauw, B., and Sandri, M. (2013). Mechanismen die de skeletspiergroei en atrofie reguleren. FEBS J. 280, 4294-4314. doi: 10.1111 / febs.12253
PubMed Abstract / CrossRef Full Text / Google Scholar
Schmalbruch, H., and Hellhammer, U. (1977). Het aantal kernen in volwassen rattenspieren met speciale verwijzing naar satellietcellen. Anat. Rec. 189, 169–175. doi: 10.1002 / ar.1091890204
PubMed Abstract / CrossRef Full Text / Google Scholar
Schwartz, L. M. (2008). Atrofie en geprogrammeerde celdood van skeletspieren. Celdood Verschilt. 15, 1163–1169. doi: 10.1038 / cdd.2008.68
PubMed Abstract / CrossRef Full Text / Google Scholar
Schwartz, L. M., Brown, C., McLaughlin, K., Smith, W., and Bigelow, C. (2016). Het myonucleaire domein wordt niet gehandhaafd in skeletspieren tijdens atrofie of geprogrammeerde celdood. Is. J. Phys. Celfysiol. 311, C607–C615. doi: 10.1152/ajpcell.00176.2016
PubMed Abstract / CrossRef Full Text / Google Scholar
Schwartz, L. M., and Ruff, R. L. (2002). Veranderingen in contractiele eigenschappen van skeletspieren tijdens ontwikkelings geprogrammeerde atrofie en dood. Is. J. Phys. Celfysiol. 282, C1270-C1277. doi: 10.1152/ajpcell.01275.2000
CrossRef Full Text / Google Scholar
Schwartz, L. M., Smith, S. W., Jones, M. E., and Osborne, B. A. (1993). Komen alle geprogrammeerde celdood door apoptose voor? Proc. Natl. Acad. Sci. U. S. A. 90, 980-984.
Google Scholar
Schwartz, L. M., and Truman, J. W. (1983). Hormonale controle van de snelheid van metamorfe ontwikkeling in de tabak hoornworm Manduca sexta. Dev. Biol. 99, 103–114. doi: 10.1016/0012-1606(83)90257-9
PubMed Abstract / CrossRef Full Text / Google Scholar
Schwartz, L. M., and Truman, J. W. (1982). Peptide en steroid regulatie van spierdegeneratie in een insect. Wetenschap 215, 1420-1421. doi: 10.1126 / wetenschap.6278594
PubMed Abstract / CrossRef Full Text/Google Scholar
Smith, H. K., Maxwell, L., Martyn, J. A., And Bass, J. J. (2000). Nucleaire DNA fragmentatie en morfologische veranderingen in volwassen konijnen skeletspieren na korte termijn immobilisatie. Cell Tissue Res. 302, 235-241. doi: 10.1007/s004410000280
PubMed Abstract | CrossRef Full Text/Google Scholar
Smith, M. I., Huang, Y. Y., and Deshmukh, M. (2009). Skeletspierdifferentiatie roept endogeen XIAP op om de apoptotische route te beperken. PLoS ONE 4: e5097. doi: 10.1371 / journal.pone.0005097
CrossRef Full Text / Google Scholar
Staron, R. S., Leonardi, M. J., Karapondo, D. L., Malicky, E. S., Falkel, J. E., Hagerman, F. C., et al. (1991). Kracht-en skeletspieraanpassingen bij vrouwen met zware weerstand na detraining en hertraining. J. Appl. Fysiol. 70, 631-640.
Google Scholar
Strassburger, E. (1893). Het werkingsgebied van de kernen en de celgrootte. Histol Beitr. 5, 97-124.
Google Scholar
Strasser, H., Tiefenthaler, M., Steinlechner, M., Eder, I., Bartsch, G., and Konwalinka, G. (2000). Leeftijd afhankelijke apoptose en verlies van rhabdosphinctercellen. J. Urol. 164, 1781–1785. doi: 10.1016 / S0022-5347 (05)67106-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Tedesco, F. S., Dellavalle, A., Diaz-Manera, J., Messina, G., and Cossu, G. (2010). Herstel van skeletspieren: regeneratief potentieel van stamcellen van skeletspieren. J. Clin. Investeren. 120, 11–19. doi: 10.1172 / JCI40373
PubMed Abstract / CrossRef Full Text / Google Scholar
Wada, K. I., Takahashi, H., Katsuta, S., and Soya, H. (2002). Geen afname van myonucleair aantal na langdurige denervatie bij volwassen muizen. Is. J. Phys. Celfysiol. 283, C484–C488. doi: 10.1152/ajpcell.00025.2002
CrossRef Full Text / Google Scholar
Winje, I. M., Sheng, X., Hansson, K. A., Solbrå, A., Tennøe, S., Saatcioglu, F., et al. (2018a). Cachexie veroorzaakt bij muizen geen verlies van myonuclei of spiervezels tijdens xenotransplantaatkanker. Acta Physiol. e13204. doi: 10.1111 / apha.13204
CrossRef Full Text / Google Scholar
Winje, I. M., Bengtsen, M., Eftestol, E., Juvkam, I., Bruusgaard, J. C., and Gundersen, K. (2018b). Specifieke etikettering van myonuclei door een antilichaam tegen pericentriolair materiaal 1 op skeletspierweefselsecties. Acta Physiol. 223: e13034. doi: 10.1111 / apha.13034
CrossRef Full Text / Google Scholar
Woo, J. (2017). Sarcopenia. Clin. Geriatr. Med. 33, 305–314. doi: 10.1016 / j. cger.2010.02.012
CrossRef Full Text / Google Scholar
Xiao, R., Ferry, A. L., and Dupont-Versteegden, E. E. (2011). Celdood-resistentie van gedifferentieerde myotubes wordt geassocieerd met verbeterde anti-apoptotische mechanismen in vergelijking met myoblasten. Apoptose 16, 221-234. doi: 10.1007 / s10495-010-0566-9
PubMed Abstract / CrossRef Full Text/Google Scholar
Yang, D., Morris, S. F., and Sigurdson, L. (1998). De sartorius spier: anatomische overwegingen voor reconstructieve chirurgen. Brigadier Radiol. Anat. 20, 307–310. doi: 10.1007
PubMed Abstract / CrossRef Full Text / Google Scholar
Zhong, H., Roy, R. R., Siengthai, B., and Edgerton, V. R. (2005). Effecten van inactiviteit op vezel grootte en myonucleaire aantal in rat soleus spier. J. Appl. Fysiol. 99, 1494–1499. doi: 10.1152 / japplphysiol.00394.2005
CrossRef Full Text / Google Scholar
Zhou, X., Wang, J. L., Lu, J., Song, Y., Kwak, K. S., Jiao, Q., et al. (2010). Omkering van kanker cachexie en spier die door ActRIIB antagonisme verspillen leidt tot verlengde overleving. Cel 142, 531-543. doi: 10.1016 / j.cel.2010.07.011
PubMed Abstract / CrossRef Full Text / Google Scholar
Ziaaldini, M. M., Marzetti, E., Picca, A., and Murlasits, Z. (2017). Biochemische pathways of sarcopenia and their modulation by physical exercise: a narrative review. Voorkant. Med. 4:167. doi: 10.3389 / fmed.2017.00167
CrossRef Full Text / Google Scholar