bevruchting in planten vindt plaats wanneer haploïde gameten elkaar ontmoeten om een diploïde zygote te creëren, die zich ontwikkelt tot een embryo. Bij gymnospermen (coniferen) en angiospermen (bloeiende planten), de ontmoeting van de gameten gebeurt op de volgende manier: mannelijke gameten zijn ingesloten in stuifmeelkorrels en worden door de wind of insecten naar de vrouwelijke voortplantingsorganen. Het eindproduct van bevruchting–het embryo–zit in een zaadje. Om deze reden worden deze twee soorten tracheofyten zaadplanten genoemd. (Zie Plantstructuren en functies, het zaad.)
Gymnosperm fertilisatie
de vrouwelijke gametofyt bevat verschillende archegonia, waar de eicellen ontstaan en zich ontwikkelen. De gametofyt zelf is omgeven door lagen van sporangia en integument; al deze elementen vormen een ovule, die wordt gevonden op het oppervlak van een vrouwelijke kegel. Bevruchting vindt plaats wanneer pollengrains (mannelijke gametofyten) worden gedragen door de wind naar het open einde van een eicel, die de eieren bevat,of vrouwelijke gametofyte. Daar ontwikkelt de stuifmeelkorrel een uitgroei, een stuifmeelbuis genaamd, die tot in de eicel in een van de archegonia doordringt. De zaadcellen in de pollentube wedijveren dan om het ei te bevruchten. Zodra de bevruchting heeft plaatsgevonden, ontwikkelt het embryo zich binnen defemale gametophyte, en de eicel wordt het zaad, compleet met een voedselbron (het gametophyte weefsel)en een zaad vacht (het integument). Dit embryo, dat uiteindelijk een nieuwe sporofyt wordt, bestaat uit twee embryonale bladeren, het epicotyl en hypocotyl.
Angiosperm fertilisatie
het vrouwelijke voortplantingsorgaan van angiospermen is de stamper, gelegen in het midden van de bloem. Net als bij gymnospermen is de mannelijke gametofyt de stuifmeelkorrel. Om bevruchting in de meeste bloeiende planten voor te komen, moeten insecten of andere dieren het stuifmeel naar de stamper transporteren. Een belangrijke onderscheidende eigenschap van angiosperms is de praktijk van dubbele bevruchting.
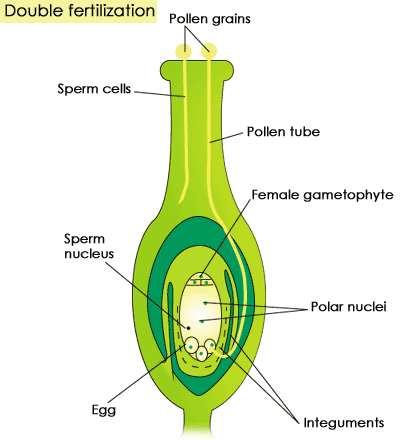
een angiosperm-ovule bevat een eicel en een diploïde fusiekern, die ontstaat door het samenvoegen van twee polaire kernen in de ovule. Wanneer een stuifmeelkorrel in contact komt met het stigma, of de bovenkant van de stamper, stuurt het een stuifmeelbuis naar beneden in de eierstok aan de basis van de stamper. Als de stuifmeelbuis de eicel binnendringt, geeft het twee spermacellen af. De ene fuseert met het ei om een diploïde zygote te creëren, terwijl de andere zich met de fusiekern verbindt om een drievoudige kern te vormen. Deze drievoudige kern verandert in een endosperm, dat het zich ontwikkelende embryo voedt (het vullen van de rol van gametofyt weefsel in het gymnosperm zaad). Net als bij gymnospermen wordt de eicel een zaadje, dat het embryo en het endosperm omhult in een zaadlaag. Maar in tegenstelling tot gymnosperms, in angiosperms ontwikkelt de eierstok die de eierstokken bevatten zich tot een vrucht na bevruchting. De vrucht geeft de embryo ‘ s het dubbele voordeel van extra bescherming tegen uitdroging en verhoogde verspreiding, omdat het wordt gegeten door verreikende dieren die vervolgens de zaden uitscheiden. (Voor een volledige bespreking van de delen van de bloem die bijdragen aan de voortplanting, zie Plantenstructuren, bloemen .)
om bevruchting mogelijk te maken, moeten angiospermen zelfbestuiven, waarbij een bepaalde plant zichzelf bevrucht, of kruisbestuiven, waarbij een plant door een andere van dezelfde soort wordt bevrucht. Kruisbestuiving produceert over het algemeen veel krachtigere planten, en wordt aangemoedigd door differentiële ontwikkeling van de mannelijke en vrouwelijke gametofyten op een bloem, of door de positionering van deze gametofyten zodat zelfbestuiving moeilijk is.