aan het begin van de negentiende eeuw merkte von Reckingausen op dat bloedvaten bedekt waren met een vel cellen. Deze eencellige dikke laag wordt samengesteld uit endothelial cellen die de interne oppervlakte van bloed en lymfevaten lijn. Bij mensen wordt geschat dat het totale oppervlak van de endotheliale laag ongeveer 35 m2 is en dat er ongeveer 1 tot 1,6 103 endotheliale cellen zijn. De totale lengte van de slagaders, aders en haarvaten is ongeveer 90000 km (meer dan twee keer reizen over de hele wereld). Endothelial cellen zijn vlak en tussen elkaar verbonden door celverbindingen. Hun belangrijkste functie is als tussenpersonen tussen het bloed en de andere weefsels, en dragen bij aan bloedeigenschappen, zowel in normale als pathologische weefsels. Ze zijn ook belangrijk voor de uitwisseling van immuuncellen tussen het bloed en weefsels.
1. Morfologie
endotheelcellen zijn zeer afgeplatte cellen, zozeer dat hun kern de hogere structuur van de cel is, zelfs als de kern ook afgeplat is. De cellulaire vorm is aangepast aan het kanaal de endotheliale cel is Voering (figuur 1, 2, en 3). In de meer smalle haarvaten, endotheelcellen kunnen uitbreiden hun cytoplasma alle omtrek van het bloedvat, zodat het kanaal is een rij van endotheelcellen. In grotere kanalen, zoals slagaders en aders, zijn veel endotheelcellen nodig voor het bedekken van de totale omtrek van het kanaal.
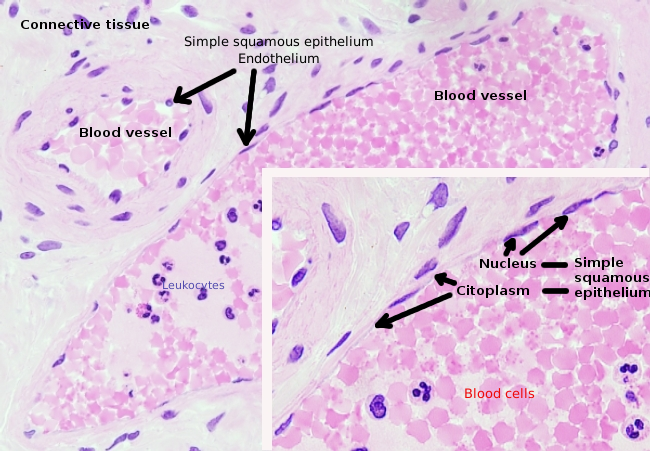
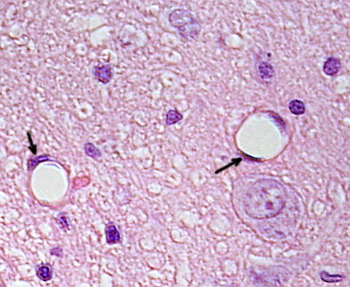
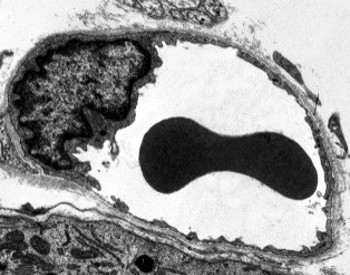
Perinucleair cytoplasma bevat de meeste mitochondriën en andere organellen, zoals Golgi-apparatuur, terwijl perifeer cytoplasma dunner is en weinig organellen heeft, hoewel endoplasmatisch reticulum dicht bij het plasmamembraan kan worden gevonden. Endotheelcellen hebben veel blaasjes, de meeste zijn endocytische blaasjes.
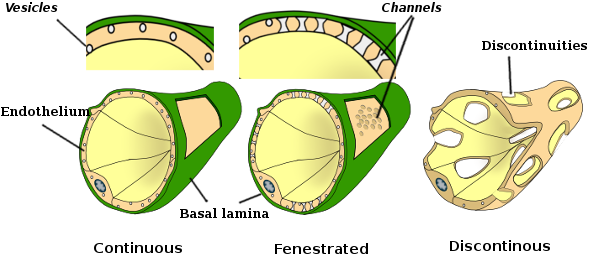
in sommige regio ‘ s, zoals lever, nierschors en endocriene klieren, vertonen endotheelcellen van haarvaten poriën of kleine passages, bekend als fenestratrions. Zij zijn wegen van ongeveer 60 tot 70 nm in diameter, met een smalle passage van ongeveer 5 nm, die direct het bloed en lymfe met de andere weefsels communiceren. Haarvaten die deze types van endothelial cellen hebben staan bekend als fenestrated haarvaten. Poriën worden verdeeld in groepen en hun dichtheid is afhankelijk van het type endotheel.
endotheelcellen worden van het surrondingweefsel gescheiden door een laag extracellulaire matrix die basaal lamina wordt genoemd. Endotheelcellen dragen bij aan deze laag met de eiwitten laminine, fibronectine en collageen type II, IV en V.
bron en proliferatie
Vasculogenese is de vorming van nieuwe endotheelcellen in weefsels zonder reeds bestaande bloedvaten. Het gebeurt in het embryo. Bij volwassenen en ook bij embryo ‘ s worden nieuwe bloedvaten, en dus nieuwe endotheelcellen, gegenereerd uit reeds aanwezige bloedvaten. Dit proces staat bekend als angiogenese. Nieuwe endotheelcellen worden geproduceerd door het vertakken of delen van bloed-en lymfevaten. De angiogenese is gemeenschappelijk in het kweken van weefsels of die onder het zware remodelleren, zoals de baarmoeder in zoogdiervrouwen, of in pathologieën zoals tumors, ontsteking en wonden.
Vasculogenese
eerste endotheelcellen van zoogdieren worden gegenereerd na gastrulatie tijdens de ontwikkeling van het embryo. Zij onderscheiden van cellen genoemd angioblasten, vormen groepen, en worden georganiseerd in korte bloedvaten. Dit proces gebeurt eerst in de vitelin zak van het embryo. Deze eerste bloedvaten groeien en verbinden zich tussen elkaar om een netwerk te vormen. Later rekruteren ze fibroblasten en spiercellen. Dit aanvankelijke netwerk ondergaat een ononderbroken het remodelleren proces tijdens de volgende ontwikkelingsstadia. In embryo ‘ s, zijn alle endothelial cellen aanvankelijk gelijkaardig, maar zij worden later gedifferentieerd in ader, slagader, capillaire en lymfatische vaatendotheel. Verscheidene molecules zoals FGF2, bmp4, IHH en VEGF veroorzaken gescheiden differentiatiewegen. De inkepingsinductor schijnt zeer belangrijk te zijn omdat het tot slagader endotheel leidt, terwijl zijn remming tot ader endotheel leidt. Lymfatisch endotheel wordt gevormd uit het endotheel van de hoofdader en heeft de expressie van COUPF-II en SOX18 nodig. Daarnaast wordt endotheel gedifferentieerd naar het orgaan waarin het zich bevindt. Bijvoorbeeld, vormt het endotheel een strak verzegelde laag in de hersenen om de bloed-hersenenbarrière te vormen, terwijl het van losjes georganiseerd in de lever wordt fenestrated om de uitwisseling van molecules met het bloed te bevorderen.
omdat endotheliale cellen zich organiseren in arteriële en veneuze kanalen, geven sommige endotheliale cellen aanleiding tot hematopoëtische cellen door een differentiatieproces dat bekend staat als endothelial-hematopoëtische transitie (EHT). Dit proces gebeurt ook in sommige volwassen weefsels zoals tijdens de vorming van placenta en andere weefsels tijdens de zwangerschap.
angiogenese
angiogenese is de vorming van nieuwe bloedvaten en endotheel door het vertakken of delen van reeds bestaande bloedvaten. Bij volwassenen is de proliferatie van endotheelcellen zeer laag. Bijvoorbeeld, in zoogdieren, kan een endothelial cel één keer per maand verdelen of verscheidene jaren wachten. Het betekent dat angiogenese zeldzaam is in weefsels onder normale fysiologie, met uitzondering van vrouwelijke voortplantingsorganen. Nochtans, kan het in pathologische processen zoals weefselschade en tumors worden geactiveerd. Sommige stoffen veroorzaken angiogenese, zoals endotheliale groeifactor (EGF), zure EGF, fibroblastgroeifactor (FGF), transformerende groeifactor (TGF) en prostaglandinen. De angiogenese begint wanneer de angiogenic substanties uit omringende weefsels worden vrijgegeven. De endothelial cel van een dichtbij bloedvat, gewoonlijk van een venule, begint aan de angiogenic signaalbron te migreren, die de andere endothelial cellen slepen, die begint te verspreiden en een nieuw schip vormen. Deze endothelial cellen verliezen tijdelijk hun strakke verbindingen resulterend in het blootleggen van het basale membraan, dat snel door enzymen wordt verteerd die van de endothelial cellen worden vrijgegeven. Sommige endothelial cellen gaan door het verteerde basale membraan terwijl de proliferatie doorgaat. Al dit proces maakt de verlenging en vertakking van het nieuwe bloedvat mogelijk totdat het vasculogenese signaal verdwijnt. De uiteinden van de bloedvaten versmelten met andere bloedvaten om gesloten circuits te vormen en de flux van bloed toe te staan zonder dode eindes.
regio ‘ s met endotheliale celvoorlopercellen zijn gevonden in de dorsale aorta en het endocardium. Mesenchymale cellen kunnen worden onderscheiden van endocardium endothelial cellen en vormen de tricuspidalisklep en sommige fibroblasten van het hart, maar niet cardiomiocytes. De Endothelial cellen zijn zo gevoelig aan externe signalen dat het houden van de endotheliumintegriteit een actief proces is, dat door andere signalen zoals FGF wordt bemiddeld. Het actieve proces is niet alleen voor integriteit, maar ook om het cellulaire fenotype te behouden. Wanneer endothelial cellen geen juiste signalen ontvangen sterven zij door apoptosis of worden mesenchymal cellen die overvloedige extracellulaire matrijs kunnen samenstellen. De overgang van endothelial aan mesenchymal cel is achter sommige ziekten zoals arteriosclerose en vezelige miocarditis.
functie
het idee dat het endotheel slechts een passieve voering van lymfatische en bloedvaten is, moet worden veranderd. Endotheel functies zijn gevarieerd en essentieel voor het organisme. Het is inderdaad een fysieke structuur van cardiovasculaire en lymfekanalen, maar het regelt ook de uitwisseling van moleculen tussen het lumen van het vat en de omliggende weefsels en beïnvloedt de fysiologische eigenschappen van het bloed. Naast een fysieke barrière ontwikkelen endotheelcellen secretorische, metabolische en immuunfuncties. Ze veranderen hun fysiologische gedrag beïnvloed door moleculen zoals groeifactoren, coagulantia en anticoagulantia, low density lipoproteïnen, stikstofmonoxide, serotonine, enkefaline, en vele anderen. Endotheelcellen hebben receptoren voor al deze stoffen.
barrière
endotheelcellen vormen een laag die gewoonlijk fungeert als een barrière tussen het bloed en de omliggende weefsels. De samenhang tussen endotheelcellen wordt vervuld door celverbindingen, zoals tight junctions en adherent junctions. Gap junctions zijn ook waargenomen, hoewel hun belangrijkste functie de communicatie tussen aangrenzende cellen is. De Endothelial cellen kunnen deze adhesie moduleren en de permeabiliteit van de barrière veranderen, die niet alleen aan molecules maar ook aan cellen kan beà nvloeden die het endotheel kruisen. Echter, in sommige organen zoals de lever, endotheelcellen zijn veel losser verpakt en laat veel vrije ruimte, zodat endotheel nauwelijks kan worden beschouwd als een barrière.
Transmissieelektronenmicroscopiebeelden tonen veel blaasjes in het cytoplasma van endotheelcellen. Ze worden verondersteld betrokken te zijn bij intracellulair transport tussen de apicale (gericht op het bloed) en basolaterale membranen (gericht op de basale lamina). Dit type transport staat bekend als transcytose. Het is interessant dat transcytose blaasjes overvloediger aanwezig zijn in de endotheelcellen van haarvaten dan in die van grotere bloedvaten. Het geeft aan dat de endotheelcellen in haarvaten een intensere uitwisseling van moleculen tussen bloed en weefsels hebben, terwijl ze in grote diameter vaten voornamelijk betrokken zijn bij het uitvoeren van het bloed. Sommige endothelial cellen, als fenestrated bekend, hebben poriën of zeer kleine kanalen die direct het bloed met de omringende weefsels verbinden, waardoor sommige kleine grootte molecules om het endotheel te kruisen zonder het cytoplasma van om het even welke endothelial cellen in te gaan (Figuur 4). Tenslotte zijn er organen met zeer permeabel endotheel. In de lever, sinusoïden zijn bloedvaten waar endothelial cellen vrije ruimte tussen elkaar verlaten, en een functie als barrière nauwelijks bestaat.
veel celtypen reizen in het bloed naar hun doelorganen in het lichaam. Daar passeren ze het endotheel van de bloedvaten, meestal op het niveau van post-capillaire aderen. Het betekent dat endothelial cellen de celverbindingen moeten wijzigen om cellen door de endotheliumlaag te laten gaan. Leukocyten verlaten het bloedvat door aan specifieke molecules van het apicale membraan van de endothelial cellen te erkennen en te verankeren. Selectins, integrins en immunoglobulins zijn verantwoordelijk voor de erkenning en adhesie van leukocyten aan de endothelial laag. Selectins beginnen het verankeren van leukocyten, die over het endothelial oppervlak rollen. Deze aanvankelijke hechting is zwak en omkeerbaar.
veel leukocyten verlaten het bloed tijdens ontstekingsprocessen om zich naar de aangetaste weefsels te verplaatsen. Chemokinen zijn leukocyten-aantrekkelijke moleculen die worden vrijgegeven door beschadigde weefsels en gekoppeld aan de glycocalix van endotheelcellen. De leukocyten die over de endothelial oppervlakte rollen die chemokines hebben worden geactiveerd, wordt de cel-celadhesie sterker, en de leukocyt blijft in de plaats. De activering van de leukocyt leidt tot de activering van zijn integrins, die immunoglobulins van de endothelial cel erkennen. Deze immunoglobulinen worden uitgedrukt in de endothelial celmembranen na de activering van de cel door chemokines. De cel-cel adhesie verhoogt de calciumconcentratie in endothelial cellen en leidt tot desorganisatie van de celverbindingen en cytoplasma terugtrekking. Op deze manier, kunnen leukocyten naar de grens van endothelial cellen bewegen en endothelium kruisen. Adhesiemoleculen zijn ook betrokken bij deze beweging van leukocyten.
bloedeigenschappen
de endotheliale functies zijn complexer dan alleen de controle van de moleculen en cellen die door het endotheel gaan. Endothelial cellen zijn ook betrokken bij bloeddruk, coagulatie, en sommige anderen bloedeigenschappen. men denkt dat de primitieve bloedsomloop 600 miljoen jaar geleden bij ongewervelde dieren is ontstaan, maar er ontbrak endotheel. Endotheliale cellen verschenen 100 miljoen jaar geleden en leverden bloed met een meer laminaire flux (niet turbulent) en daardoor een efficiëntere gasuitwisseling.
endotheelcellen moduleren de bloeddruk door stoffen vrij te geven die inwerken op de gladde spieren van bloedvaten. Ze geven stikstofmonoxide (NO) en prostacycline af, die de gladde spieren van de bloedvaten ontspannen. Zij geven ook endotheline en de plaatjesactivatorfactor vrij, beide verminderend de bloedvatdiameter. Stikstofmonoxide wordt constitutief vrijgegeven en zorgt voor een goede spierspanning, remt bloedplaatjesaggregatie en leukocytenadhesie. Endotheline is een sterke vasoconstrictor. Hoe endothelial cellen beslissen welke molecule moet worden vrijgegeven is nog niet duidelijk, maar mechanorreceptoren in hun apicale membranen die in staat zijn om de flux eigenschappen van het bloed kunnen plausibel zijn.
onder normale omstandigheden geven endotheelcellen moleculen af in het bloed die helpen een goede vloeibaarheid te behouden. Ze werken op twee niveaus: vloeibaarheid (anticoagulantia) en het voorkomen van bloedplaatjesaggregatie (antitrombotica). Eiwitten C en S zijn belangrijke moleculen die de bloedstroom beïnvloeden. C-eiwit, dat een complex vormt met s-eiwit, inactiveert de stollingsfactoren VIIIa en Va. S eiwit wordt gesynthetiseerd door endothelial cellen. Bovendien bevat het endotheliale glycocalix een glycosaminoglycaan vergelijkbaar met heparine dat trombine kan inactiveren.
in verband met hun antistollingsfunctie geven endotheelcellen stikstofmonoxide en prostacycline af, waardoor zowel het cyclisch AMP in bloedplaatjes stijgt als de bloedplaatjesaggregatie moeilijker wordt. Deze twee moleculen komen continu vrij in het bloed. Endothelial cellen hebben ectonucleases in hun apicale membranen, die ATP en ADP, zowel sterke promotors van plaatjesaggregatie verwijderen. Bovendien geven endotheliale cellen een activatormolecuul af dat plaminogen omzet in plasmine, wat het verwijderen van trombi bevordert.
al deze moleculaire routes kunnen veranderen wanneer endotheelcellen bepaalde signalen ontvangen of wanneer weefsel beschadigd is, wat leidt tot bloedstolling en bloedplaatjesaggregatie. In deze omstandigheden worden endotheelcellen dan actieve deelnemers aan de coagulatie en trombose.
immuunafweer
endotheelcellen spelen een belangrijke rol in de immuunafweer en zijn betrokken bij twee mechanismen: de presentatie van antigenen tegen T-lymfocyten en het werven van immuuncellen. Samen met macrofagen, kunnen endothelial cellen antigenen aan T-lymfocyten voorstellen omdat zij constitutief MHC-I (major histocompatibility complex) uitdrukken en kunnen worden veroorzaakt om MHC-II uit te drukken, beide noodzakelijk voor antigeenpresentatie. Endothelial cellen kunnen immune geheugen activeren, maar geen nieuwe T-lymfocyten. Er is bidirectionele activering tussen endothelial cel en T-lymfocyten, zodat endothelial cel molecules voor het aantrekken van ontstekingscellen vrijgeven en adhesiemolecules voor het verankeren van bloedleucocyten uitdrukken.
Bibliografie
Cines BD, Pollak ES, Buck CA, Loscalzo J, Zimmerman GA, Mcever RP, Pober JS, Wick TM, Konkle BA, Schwartz BS, Barnathan ES, McCrae KR, Hug BA, Schmidt A-M, Stern DM 1998. Endotheelcellen in de fysiologie en in de pathofysiologievan vaataandoeningen. The journal of the american society of hematology. 91:3527-3561 ![]()
Fajardo LF. 1988. De complexiteit van endotheelcellen. American journal of clinical pathology. 92:241-250.
Michiels C. 2003. Endotheliale celfuncties. Journal of cellular physiology. 196: 430-443 ![]()