Introduksjon
Internasjonal bevaringspolitikk anerkjenner biologisk mangfold på tre nivåer, økosystem, arter og genetisk, og at ledelsen bør sikte på å beholde alle tre (Konvensjon Om Biologisk Mangfold, 2007). Dette gjenspeiles tydelig i Konvensjonen Om Biologisk Mangfold, Aichi Biodiversity Targets, avtalt i 2010 (https://www.cbd.int/sp/targets/), der det er spesifikk referanse i mål og mål, ikke bare til økosystemer og arter, men også til genetisk mangfold. Likevel er dagens tilnærminger til bevaring av biologisk mangfold i stor grad basert på geografiske områder, økosystemer, økologiske samfunn og arter, med mindre oppmerksomhet på genetisk mangfold og artspopulasjonskontinuumet. Faktisk fokuserer flertallet av bevaringsutøvere på arter som metrisk av biologisk mangfold, med noe hensyn til intraspesifikke enheter innenfor intensivt forvaltede truede arter. I denne sammenheng er det et naturlig ønske om å benytte enkle kategoriseringer-navngitte arter og andre navngitte taxa( dvs. underarter), og noen ganger bevaringsenheter innenfor arter. Spesielt arter gir et felles mål for biologisk mangfold som støtter mye vitenskapelig og ledelsesarbeid (Mace, 2004; Wilson, 2017). Som en grunnleggende enhet for bevaring brukes arter ofte til å kvantifisere biologisk mangfold verdi gjennom lister over arter innenfor verneområder, identifisering av truede arter innenfor jurisdiksjoner, og som grunnlag for biosikkerhetsprosedyrer. I tillegg er arter vanligvis de enhetene som vi kommuniserer bevaringsspørsmål til publikum.
denne impulsen til å bare identifisere diskrete enheter i naturen står i motsetning til flere relaterte realiteter. For det første er spesiering ikke øyeblikkelig. Snarere er det vanligvis en langvarig prosess (Dynesius Og Jansson, 2014; Roux et al., 2016), fremgang fra isolerte populasjoner, kanskje utsatt for forskjellige utvalgstrykk, til entydige arter over lange tidsskalaer, med hyppige reverseringer forårsaket av miljøendringer eller enkel genetisk sammenslåing som områder kobles til igjen (Figur 1; Seehausen et al., 2008; Rosenblum et al., 2012). For det andre er de fleste anerkjente arter ikke genetisk ensartede. Divergens (drift og langvarig isolasjon) og retikulasjon (genflow, hybridisering, introgression og rekombinasjon) danner effektivt geografisk og genomisk variasjon innen arter (Cutter og Payseur, 2013; Edwards et al., 2016) og noen arter kan struktureres i lange isolerte populasjoner, hvorav noen kan garantere anerkjennelse som fulle, men kanskje morfologisk kryptiske arter (Bickford et al., 2007; Jorger Og Schrö, 2013). Tredje, så lenge anerkjent, navngitte arter er ikke ekvivalente med hensyn til hvor mye evolusjonær arv de representerer-enkelt sagt, en platypus (som representant for en gammel divergert avstamning) kan anses å være verdt mer enn en nylig divergert gnager (Isaac et al., 2007). For det fjerde er artsbetegnelser ikke kastet i stein, men de er hypoteser som skal testes når nye bevis blir tilgjengelige (Hey et al., 2003). Hvert av disse problemene skaper kompleksitet og tvetydighet i å avgrense arter generelt, og tildele bevaringsoppmerksomhet til dem eller deres komponentpopulasjoner. I tillegg er avgrensningen av arter også gjenstand for forskjellige artsbegreper og måter å diagnostisere dem på, noe som har forårsaket stor bestyrtelse blant bevaringsbiologer (Mace, 2004; Frankham et al.(2012; Garnett Og Christidis, 2017).
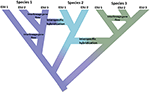
Figur 1. Spesiering som et kontinuum som går fra isolerte populasjoner til entydige arter vist ved skiftet i farge med fargene som representerer forskjellige arter. Noen linjer som oppstår på grunn av befolkningsisolasjon utvikler seg til sanne arter med de indre noder av treet som representerer initiering av spesiasjonsprosessen. Andre linjer utvikler seg ikke til sanne arter, men divergerer over tid, noe som indikerer at artene ikke er genetisk ensartede. Disse linjene er vist Som Evolutionarily Significant Units (Esu). Inter-lineage genflyt er indikert For Art 1 Og Art 3. I dette tilfellet kan evolusjonær uavhengighet være forbigående fordi to linjer som tilhører samme art ikke er reproduktivt isolert. Mens de forblir som forskjellige linjer (arter), kan interspesifikk hybridisering forekomme med begrenset genutveksling (skifte i farge) mellom arter.Implementering av effektive bevaringsstrategier som tar sikte på å beskytte både arter og genetisk mangfold, vil ha nytte av både et mer konsistent syn på hvordan vi avgrenser arter og andre enheter på tvers av populasjonsartkontinuumet, og hvordan disse enhetene tolkes i forskjellig lovgivning globalt. Nye evner fra genomics gir både muligheter og utfordringer for å nå disse målene. I det følgende vurderer vi: (i) hvordan genomskalaanalyser kan avstemmes med en konservativ tilnærming til avgrensning og navngivning av arter, samtidig som man identifiserer viktige komponenter i genetisk mangfold innen arter, (ii) hvordan eksisterende bevaringslovgivning på flere kontinenter, som definerer «arter» for å inkludere et bredt spekter av infra-spesifikke kategorier, kan brukes gitt den høye oppløsningen som tilbys av genomskjermer, og (iii) hvordan innsikten i artsprosesser og strukturering av variasjon fra genomikk samsvarer med lovgivning og deretter inn i ledelse. Vi foreslår deretter noen måter fremover i å tilpasse bevaringsenheter med lovgivningsmessige tilnærminger for å beskytte hele spekteret av mangfold fra populasjoner gjennom arter.
Genomikk, Artsdannelse, Og Taksonomi
som mange felt av biologisk vitenskap, systematikk, praksisen med å navngi taxa, og forstå deres relasjoner og hvordan de danner, blir betydelig avansert av genomikk og statistiske modeller. Enten gjennom hele genomsekvensering (F.eks., 2017) eller genomprøvetaking (SNPs, target capture-sequencing; Jones and Good, 2016; Leache and Oaks, 2017), har vi nå enestående oppløsning av mønstre av genetisk mangfold (Funk et al., 2012). Sammen med stadig kraftigere statistiske modeller for å utlede historier om genomisk og artsdivergens, gir disse dataene ny innsikt i evolusjonære prosesser som genererer arter og genetisk variasjon.
Anvendelse av disse nye verktøyene genererer to motstridende innsikt. På den ene siden avslører mange studier svært divergerende genetiske populasjoner innen navngitte arter, såkalte kryptiske arter (Bickford et al., 2007; Jorger Og Schrö, 2013; Strik et al., 2017). Dette er ikke noe nytt, da det bygger på en lang historie med romlig populasjonsgenetikk og molekylær fylogeografi som har vært en produktiv innsats i tre tiår (Avise et al., 1987). Hva genomikk bringer til dette arbeidet er både økt oppløsning av befolkningsstruktur og data som er tilstrekkelig til å statistisk utlede befolkningshistorier og teste alternative modeller av divergens (Degnan Og Rosenberg, 2009; Sukumaran og Knowles, 2017). På den annen side avslører genomskalaanalyser også at genetisk utveksling (introgression) blant dyrearter er mer vanlig enn tidligere antatt, både under og etter artsdannelse, og kan til og med drive nye adaptive strålinger (Mallet, 2007; Rieseberg og Willis, 2007; Arnold Og Kunte, 2017) Figur 1. Samlet sett understreker disse innsiktene at spesiering er langvarig (Roux et al., 2016) og avsløre de motsatte kreftene som opererer gjennom fylogeografi-fylogenetikkkontinuumet (Edwards et al., 2016).
Hvordan henger alt dette sammen med taksonomi og bevaringsmål? Det er sterk og forsvarlig bekymring for at den økte oppløsningen som gis av genomskjermer, kan føre til voldsom oversplitting av arter (Isaac et al., 2004), potensielt begrense ledelsesfleksibilitet og sende små genetisk divergerende populasjoner til innavl og eventuell utryddelse (Frankham et al ., 2012). Omvendt, Gippoliti et al. (2018) hevder at taksonomisk treghet, noe som resulterte i tidligere manglende anerkjennelse av Mange arter Av Afrikanske hovdyr, hadde resultert i utavlsdepresjon etter blanding og manglende beskyttelse av geografisk begrenset taxa. Deres syn (og se Også Morrison et al., 2009) er at det ikke er bevis for at taksonomisk inflasjon har negative effekter på ungulat bevaring (Gippoliti et al., 2018). En relatert bekymring er at ustabil taksonomi forsinker både praksis med bevaring og offentlige oppfatninger (Garnett og Christidis, 2017). Disse bekymringene er delvis drevet av den lange løpende debatten om artsbegreper og diagnostiske metoder i systematisk biologi, særlig anvendelse av Det Fylogenetiske Artsbegrepet som hevder at arter er minimalt diagnostiserbare enheter (Cracraft, 1983). Imidlertid er flertallet av taksonomer iboende konservative, bare navngi arter når det er samsvar over flere bevislinjer (Dayrat, 2005; Padial et al., 2010; Schlick-Steiner et al., 2010: Yeates et al., 2011). Vårt syn er at arter skal representere robuste uavhengig utviklende linjer som forblir stort sett intakte når de kommer i kontakt med nære slektninger – i samsvar med Hensikten Med Evolusjonær Artsbegrepet (Wiley, 1978) og Generalisert Artsbegrep (De Queiroz, 1998, 2007). I praksis krever dette direkte bevis på reproduktiv isolasjon eller rimelige surrogater av det samme, samtidig som det tillates begrenset genetisk utveksling (Singhal et al., 2018).Genomiske data gir større kraft enn før for å identifisere differensiering og divergens innenfor og blant arter, og krever større vurdering av hvordan dette påvirker avgrensning av taksonomiske enheter. Hvor populasjoner er både sterkt genetisk og fenotypisk divergerende i forhold til allerede navngitte arter, er det et sterkt tilfelle for å navngi disse som separate arter. Hvis populasjoner er sterkt divergerende fenotypisk med liten genomisk separasjon, så er det en dom samtale om disse fenotypiske forskjellene er (i) arvelig, og (ii) sannsynlig å forårsake substansiell reproduktiv isolasjon. Genomskanninger kan løse slike situasjoner ved å teste for begrenset genflyt. Det mer utfordrende problemet er hvor morfologisk like, men genetisk divergerende populasjoner oppdages. Slike populasjoner kan lett bli diagnostisert som separate evolusjonære linjer ved å anvende statistiske avgrensningsmetoder til multilocus data (Carstens et al., 2013; Rannala, 2015), men er de flyktige populasjoner eller holdbare arter (Sukumaran Og Knowles, 2017)? Det er sikkert mulig, innenfor nomenklaturens regler, å gjenkjenne arter fra DNA-sekvenser alene, og gi sekvensbasert diagnostikk (Tautz et al., 2003; Cook et al., 2010). For eksempel, Murphy et al. (2015) avgrense og beskrive seks Australske ørkenfjær amphipod arter basert PÅ DNA-sekvensdata til tross for ingen klare morfologiske forskjeller. Dette er imidlertid et generelt forbedringsområde der systematikere må bruke begrepet «kryptiske arter»med større konsistens og strenghet (Struck et al., 2017).I situasjoner DER DNA-sekvensdata gir et potensielt nøkkeldefinerende trekk for anerkjennelse av en art, foreslår vi at det er spesielt viktig å bruke andre bevislinjer for å unngå oversplitting (Oliver et al., 2015). En tilnærming er å teste for betydelig reproduktiv isolasjon der slike populasjoner danner naturlige kontaktsoner. For eksempel, i en komparativ genomisk analyse av kontakt soner blant avstamning-par Av Carlia øgler som er fenotypisk utvisket, indekser av genetisk isolasjon var sterkt korrelert med divergens tid (Singhal Og Moritz, 2013; Singhal Og Bi, 2017) (Figur 2). De mer divergerende av disse avstamning-parene har nå blitt formelt anerkjent som separate arter, mens mer nært beslektede (ennå statistisk diagnostiserbare) linjer med bevis på genomisk omfattende genetisk utveksling på tvers av kontaktsoner var ikke (Singhal et al., 2018). Gitt det sterke empiriske forholdet mellom divergens og reproduktiv isolasjon i disse taxaene, separerte disse forfatterne ytterligere to allopatriske og kryptiske isolater (av en nært beslektet art) som hadde enda høyere genomisk divergens enn de som er avgrenset ovenfor. Utvide denne reproduktive isolasjonen informert metrisk til andre kongeneriske arter med dypt divergerende, men morfologisk lignende fylogeografiske linjer (Potter et al., 2016, 2018; Afonso Silva et al., 2017a) antyder at flere arter gjenstår å bli beskrevet, spesielt på øyer, (Figur 2, Carlai amax Og C. rufilatus), og faktisk har noen slike arter blitt beskrevet etter komplementær fenotypisk analyse (Afonso Silva et al.( 2017b). Andre linjer med nyere avvik, men fortsatt statistisk avgrenset som separate linjer ved hjelp av et stort sett med nukleare gener, kan representeres Som Evolusjonært Signifikante Enheter (Esu) innen arter.
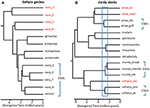
Figur 2. Fylogenomisk oppløsning av artsgrenser i øgler fra De Australske Monsoonal Tropics (AMT). I hvert tilfelle er forhold og relative divergenstider estimert fra 100 eksoner ved hjelp av multispecies coalescent implementert I StarBEAST2, men tidsskalaene mellom de to eksemplene er ikke nødvendigvis de samme. (A) Resultater for nana gruppen Av Gehyra geckos (Moritz et al., 2018). Den fylogenetiske mangfold av linjene Innenfor G. nana, som tidligere anerkjent, inkludert fire andre allerede beskrevet og morfologisk særegne arter. Etterfølgende analyser av områdeoverlapper og morfologi har ført til anerkjennelse av hver av de fylogenetisk divergerende linjene (røde taksonetiketter) som separate arter (Doughty et al., 2018). De (morfologisk uadskillelige) linjene som er igjen I G. nana som nå anerkjent, kan betraktes Som ESUs. (B) Lopende opplosning av artsgrenser i Carlia skinks fra AMT. Noen linjer har allerede blitt anerkjent som arter basert på en kombinasjon av genetisk art avgrensning og post hoc identifikasjon av diagnostiske morfologiske forskjeller (C. insularis og c. isostriacantha; Afonso Silva et al.( 2017b). For andre komplekser (Potter et al., 2016, 2018), taksonomiske revisjoner pågår. Det blå rektangelet indikerer rekkevidden av divergenstider der det er observert betydelig reproduktiv isolasjon, korrelert med divergens, i kontaktsoner mellom morfologisk kryptiske linjer av beslektede arter (Singhal og Bi, 2017). Konservativt, og følgende (Singhal et al., 2018), anerkjenner vi som kandidatarter tre genetiske linjer med avvik større enn denne empiriske terskelen. Dette ville etterlate genetisk distinkte ESUs innenfor Hver Av Carlia amax, Carlia munda og Carlia rufilatus (blå parentes). Skalaen barer representerer estimater av divergens tid i millioner av år.i ekstreme tilfeller avslører genomiske metoder at enkelt taksonomiske arter ikke bare er genetisk heterogene, men i fylogenetisk forstand også inneholder andre navngitte arter. Slike «paraphyly» kan oppstå om nylig spesiering fra en enkelt, genetisk oppdelt art (Patton og Smith, 1994), men det genererer også hypotesen om at det genetisk heterogene taxonet kan representere flere arter. Igjen, mer bevis er nødvendig. Som et tilfelle i punkt, fylogenomisk analyse Av et kompleks Av Gehyra gecko øgler fra nord-Australia avgrenset ni evolusjonære linjer, med fire allerede navngitte arter fylogenetisk innebygd blant dem (Moritz et al ., 2018; Figur 2). Senere bevis på geografisk overlapping (samtidig som fylogenetisk separasjon beholdes) og subtile morfologiske forskjeller støttet navngiving av de fire mest fylogenetisk divergerende linjene (Doughty et al., 2018). Omvendt hadde de mer nært beslektede (men statistisk separerbare) linjene tilstøtende snarere enn overlappende fordelinger, men var ikke morfologisk diagnostiserbare ,og ble derfor beholdt som en enkelt art (G. nana sensu stricto), om enn med flere Esuer, i påvente av videre undersøkelse (Figur 2). Etterfølgende genomskanninger av prøver over grensene til Disse Esuene viste at i det minste noen har betydelig genetisk utveksling, validering av den opprinnelige, konservative tilnærmingen tatt for å gjenkjenne arter.i andre tilfeller kan plantegenomiske studier, selv gjennom enkel ordinering Av SNPs, avsløre sterke og uventede avvik mellom populasjonsprøver, noe som tyder på tilstedeværelsen av kryptiske taxa som deretter kan analyseres for diagnostiske fenotypiske tegn. FOR eksempel, SNP-baserte bevis for genomisk divergens mellom populasjoner og økotypiske forskjeller I Eukalyptus salubris avdekket to distinkte molekylære linjer som opprettholdt sin egenart i et område av geografisk overlapping, med økotypisk tilpasning anses å være en viktig faktor i å minimere genflyt mellom linjene. (Steane et al., 2015; Figur 3). I Et annet tilfelle, I Australsk Pelargonium, genomisk analyse, kombinert med morfologiske og reproduktive studier, har ført til betydelige endringer i taksonomi, både splitting og lumping av taxa, spesielt der tidligere brukte morfologiske egenskaper er funnet å være labile og uharmoniske med fylogeni (Nicotra et al., 2016).
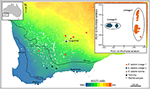
Figur 3. Genomisk analyse ved HJELP AV SNP data identifisert kryptiske linjene i den utbredte Eucalyptus salubris. Plasseringen av prøver populasjoner av hver avstamning er vist over en aridity index gradient i south west Western Australia. PCoA basert på binære genetiske avstander avledet fra 16,122 DArTseq markører, viser to forskjellige linjer. Tilpasset Fra Steane et al. (2015).ovennevnte tilfeller illustrerer den progressive hypotesetestingsmetoden som har blitt brukt i integrativ taksonomi (Padial et al., 2010; Yeates et al., 2011). Videre bruker disse tilfellene noen form for genomreduksjonssekvensering for å oppnå data av høy kvalitet fra 1000 av uavhengige gener, noe som gir mye større tillit til resultatene enn det som er tilfelle ved bruk av bare plastid loci eller noen få nukleare gener. Et eksempel på hvordan bruk av integrativ taksonomi kan dramatisk endre oppfatninger av mangfold kommer fra en kombinert genetisk, akustisk og morfologisk undersøkelse av frosker fra Madagaskar som nesten doblet det kjente artsmangfoldet (Vieites et al., 2009). Disse forfatterne foreslo en rekke vilkår som gjenspeiler tilstanden til taksonomisk oppløsning og kongruens: Bekreftede Kandidatarter, for genetisk og fenotypisk divergerende, men ennå ikke navngitt taxa; Ubekreftede Kandidatarter for genetisk divergerende taxa hvor morfologi og akustikk ennå ikke var undersøkt; Og Dypt Spesifikke Linjer for enheter kjent for å være genetisk divergerende, men fenotypisk kryptisk. Genomiske analyser er ennå ikke brukt på disse taxa, men vil sikkert informere beslutninger om spesifikke linjer vs kandidatarter.Genomiske tilnærminger, selv når de brukes konservativt, vil uten tvil føre til ytterligere splitting av morfologisk konservative taxa. Men omvendt er det klare eksempler hvor de samme tilnærmingene avslører oversplittede eller feildiagnostiserte arter (Nicotra et al., 2016). Selvfølgelig er det allerede mange molekylære studier på plante – og dyreartskomplekser basert på mindre datasett (f. eks. mikrosatellitt, mtDNA og cpDNA) som har identifisert hvor arter har blitt feildiagnostisert, eller hvor flere arter har blitt omskrevet taksonomisk, men nå anses å være en enkelt art. For eksempel, Rabosky et al. (2014) brukte mitokondrielle og nukleare genfylogenier for å revidere scincid Ctenotus inornatus-komplekset, noe som resulterte i en netto reduksjon av arter og justering av artsgrenser; de morfologiske egenskapene som tidligere ble brukt, mens det var praktisk for feltbiologer, var svært labile og i stor grad ikke klarte å matche artsgrenser som diagnostisert genetisk. I planter noen av de mest kjente eksemplene er funnet i orkideer, slik Som Den Europeiske orkide slekten Ophrys og Australian orchid slekten Caladenia, hvor molekylære studier indikerer betydelig færre enheter som ville bli anerkjent som arter, enn de som er basert på morfologiske egenskaper og pseudocopulatory pollinering syndromer (se Devey et al., 2008; Swarts et al., 2014).
Data I Genomskala, når de tolkes konservativt og ideelt med parallelle analyser av fenotyper, vil fortsette å føre til taksonomiske endringer, spesielt i morfologisk konservative taxa (Fiš et al., 2018). Men i motsetning til bekymringer uttrykt av noen (Garnett Og Christidis, 2017), anser vi at dette er den riktige jakten på evolusjonær systematikk, selv om resultatene er midlertidig ubeleilig.
Genomikk og Intraspesifikke Bevaringsenheter
Selv om det er mindre omstridt enn taksonomisk avgrensning og artsnivådebatten, men på mange måter like kompleks, har det vært den betydelige diskusjonen om definisjon og avgrensning av bevaringsenheter som viktige elementer i intraspesifikt mangfold som må vurderes i bevaringshandlinger (Ryder, 1986; Waples, 1991; Dizon et al., 1992; Moritz, 1994; Vogler og De Salle, 1994; Crandall et al., 2000; Fraser Og Bernatchez, 2001; Avise, 2004; Funk et al. 2012 (Figur 4). Gitt den forsiktige tilnærmingen til å gjenkjenne arter fra genomisk bevis som vi fortaler ovenfor (og se Også Oliver et al., 2015; Singhal et al., 2018), er det uunngåelig at noen taksonomiske arter fortsatt vil inneholde lange isolerte populasjoner i dem. Begreper Som Evolusjonært Signifikante Enheter (Esuer) eller Betegnbare Enheter (Duer) har blitt foreslått å utfylle eksisterende taksonomi (Ryder, 1986; Waples, 1991; Moritz, 1994; Mee et al., 2015). Som med arter varierer tolkninger og kriterier, men den felles hensikten er å gjenkjenne viktige elementer i intraspesifikt mangfold for bevaringshandlinger (Funk et al., 2012). Moritz (1994, 2002) foreslo en forenklet, men praktisk molekylær tilnærming til å identifisere uavhengig utviklende segmenter innen arter, den» historiske «aksen av mangfold; mens andre legger mer vekt på den «adaptive» aksen for å inkludere funksjonelt mangfold (Ryder, 1986; Crandall et al., 2000; Fraser og Bernatchez, 2001; Waples et al., 2001; Mee et al. 2015 (Figur 4). Uansett de relative fordelene ved disse alternativene, er det klart at de tekniske og analytiske verktøyene for populasjonsgenomikk nå robust kan identifisere begge komponentene av intraspesifikt mangfold(Funk et al., 2012; Hoban et al., 2013; Whitlock, 2014; Yang og Rannala, 2014).
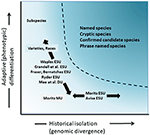
Figur 4. Separasjon av genetisk mangfold i to komponenter: adaptiv variasjon som oppstår gjennom naturlig utvalg, og analysert gjennom analyse av fenotyper og historisk isolasjon eller nøytral divergens vurdert gjennom genomisk divergens. Det mørkere skyggelagte området over kurven indikerer forhold der populasjoner sannsynligvis vil bli vurdert som separate arter under de fleste konsepter. Ulike bevaringsenhetskonsepter og infrarøde taksonomiske kategorier er vist i forhold til de to aksene av genetisk mangfold (Etter Moritz, 2002).
dette fører til spørsmålet: hvorfor ikke bare nevne ESUs som underarter? Underarter har tradisjonelt blitt definert og sett av både botanikere og zoologer som fenotypisk distinkte, allopatriske sett av populasjoner som kan intergrade inn i hverandre ved geografiske grenser (Mayr, 1942; Grant, 1981). Som sådan reflekterer de potensielt adaptiv divergens uten nødvendigvis å ha vært historisk isolert. Underarter betegnelser er mye brukt i planter, med geografiske og økologiske forskjeller den primære måten de utmerker seg på (Hamilton Og Reichard, 1992). I den grad underarter manifesterer lokal tilpasning, med eller uten pågående genflyt, foreslår vi at fenotypisk definerte underarter fortsatt skal gjenkjennes i bevaring, og ikke sammenblandet Med ESUs, i det minste Ikke ESUs som definert for å representere historisk isolerte populasjoner.på samme måte som populasjonsgenomikk gir langt større presisjon i identifisering Av Esu ved å benytte både nøytrale og adaptive loci, gir det også et middel for å utnytte et stort antall nøytrale loci for lettere å avgrense Styringsenheter (MUs) som demografisk uavhengige enheter preget av begrenset genflyt (Funk et al., 2012). Mens det meste av vårt fokus her er PÅ ESUS og analoge enheter Som DUs som kan anerkjennes som de viktigste intraspesifikke enhetene for bevaring og har blitt akseptert for beskyttelse under noen jurisdiksjoner, Har MUs også blitt identifisert som representerer demografisk uavhengige sett av populasjoner (se Palsbø et al., 2007) som kan klare å beholde den større ESU (Moritz, 1994; Funk et al., 2012). Forvaltningsenheter kan være viktige for overvåking og styring av populasjoner av arter og har også blitt referert til som «bestander», spesielt relevant for fiskeriforvaltning (Avise, 2004; Laikre et al., 2005). Definisjonen Av MUs Av Moritz (1994)som «populasjoner med betydelig divergens av allelfrekvenser ved kjernefysiske eller mitokondrie loci, uavhengig av fylogenetisk egenart av alleler» er generelt allment akseptert, selv om, som påpekt Av Palsbø et al. (2007), demografisk tilkobling er nøkkelfaktoren ikke nivået av historisk genflyt.Å Avgrense bevaringsenheter som viktige elementer i intraspesifikt mangfold har også betydelige implikasjoner for genetisk redning av små innavlede populasjoner gjennom restaurering av genflyt eller ved forsterkning, og det blir stadig tydeligere at genetisk redning må vurderes bredere hvis økt befolkningsutryddelse skal avverges (Love Stowell et al., 2017; Ralls et al., 2018). Det Ble foreslått Av Moritz (1999) at blanding Av MUs, men Ikke ESUs, kunne betraktes som en hensiktsmessig strategi for genetisk redning. Som understreket Av Frankham et al. (2012) blanding av taksonomisk anerkjente arter vil generelt ikke anses som akseptabelt av ledere, i så fall kan taksonomisk oversplitting begrense mulighetene for å redde små innavlede populasjoner. Vi foreslår at mens en strengere tilnærming til å avgrense intraspesifikke enheter som MUs kan bidra til å lindre dette problemet, er det også klart at genomisk estimering av migrasjonsrater og adaptiv divergens har potensial til å gi mye mer robust veiledning i ikke bare å definere artsgrenser og intraspesifikk variasjon, men i utformingen av genetiske redningsstrategier.
Fylogenomikk og Bevaring
Genomikk kombinert med fylogenetiske metoder kan lett løse relasjoner og evolusjonær avstand mellom taxa, samt avgrense arter. Dette oppnås best ved hjelp av arter tre metoder, snarere enn sammenkobling, som den tidligere konto for gent tree heterogenitet og unngå over-inflasjon av spisslengder på grunn av forfedrenes polymorfisme (Edwards et al., 2016; Ogilvie et al., 2016). Mens massive antall loci er ofte ikke avgjørende, inkludert flere loci forbedrer ofte oppløsning av både relasjoner og gren lengder (f.eks Blom et al., 2016). Betydningen for bevaring er todelt. For det første muliggjør fylogenetiske vurderinger vurdering av artenes eller Esusens særpreg, ikke bare om de er beskrevet. DERMED KANTEN programmet (Isaac et al., 2007) fortaler vekting av arter for bevaring basert på hvor mye unik evolusjonær historie de representerer, i tillegg til vanlige iucn-kriterier. For det andre kan grenlengder selv representere biodiversitetsverdier, uavhengig av taksonomi (Faith, 1992), som har vist verdi i å prioritere områder for bevaring (Laity et al., 2015; Rosauer et al., 2016, 2018). I denne forstand gjør vedtak av fylogenetiske beregninger ved hjelp av tiltak som fylogenetisk mangfold og fylogenetisk endemisme det mulig å markere områder med høyt evolusjonært mangfold som kan målrettes for beskyttelse, og gis høy prioritet i enhver planprosess for forbedret bevaring av biologisk mangfold. Denne tilnærmingen kan forbedre beskyttelsen av genetisk mangfold og evolusjonære prosesser for ulike grupper av organismer på tvers av en rekke geografiske skalaer (Laity et al., 2015) ta beslutninger om områder for å prioritere mer robust til vagaries av hvordan genomisk bevis er oversatt til anerkjennelse av arter eller intra spesifikke enheter.
Den Virkelige Verden-Lovgivende Definisjoner
Politikk og lovgivningsrammer for bevaring av biologisk mangfold spenner fra internasjonale konvensjoner og strategier, gjennom nasjonale strategier og lovgivning, og statlig eller regional lovgivning og strategier innen land. De viktigste internasjonale virkemidlene for biologisk bevaring er Konvensjonen om Biologisk Mangfold som er implementert gjennom Partskonvensjonen (COP), Verdensarvkonvensjonen, Den Globale Biodiversitetsstrategien, Montreal-Prosessen for Skogvern, Ramsar-Konvensjonen om Våtmarker, Konvensjonen om Trekkende Arter, Konvensjonen Om Internasjonal Handel Med Truede Arter (CITES), Den Internasjonale Naturvernunionen (IUCN) rødliste over truede arter og Fns rammekonvensjon om klimaendringer. Alle disse charterene legemliggjør prinsippet om å bevare arter og genetisk mangfold som et grunnleggende mål for bevaring av biologisk mangfold, med arter som enheten bredt definert for beskyttelse. Definisjonene av arter i henhold til lovgivning og internasjonale direktiver er imidlertid svært variable. De inkluderer taksonomisk anerkjennelse av arter og intraspesifikke taxa, men kan også inkludere genetiske/evolusjonære begreper som ofte dekkes av den juridiske definisjonen av en » art » (Tabell 1).

Tabell 1. Verdens lovgivning og direktiver som definerer «arter» for oppføring og beskyttelse.
Gitt det brede spekteret av infrarøde nivåer som kan beskyttes (Tabell 1), bør lovgivningen og tilhørende listingsprosesser i land legge til rette for bevaring av genetisk mangfold. For eksempel kan definisjonen av en «art», når man vurderer lovgivning rundt om i verden (Tabell 1), omfatte: arter, underarter, varianter og geografisk og/eller genetisk distinkte populasjoner, mens Under Sørafrikansk lovgivning kan det også omfatte kultivarer, geografiske raser, stammer og hybrider. Under de fleste lovgivning globalt omfatter en » art » enheter som reflekterer ulike infrarøde nivåer som alle er en del av en bredere artsfordeling og reflekterer i varierende grad populasjonsartens kontinuum. Dette paralleller den uunngåelige vanskeligheten som taksonomer, evolusjonære biologer og bevaringsgenetikere står overfor i å finne en mer konsistent og ensartet måte å spesifisere enheter innenfor dette kontinuumet.
Noen hevder at bekymringer om de politiske implikasjonene av arter og infrarød usikkerhet er feilplassert (Hey et al., 2003), men vi er fortsatt av den oppfatning at mens lovgivningen kan gi en mekanisme for å beskytte genetisk mangfold på en rekke nivåer, er det på biologer å gi mer klarhet og konsistens på hvordan disse nivåene skal avgrenses fra arter til individuelle populasjoner. I dag ledere og lovgivere sitter igjen med en forvirrende rekke artsbegreper og infra-spesifikke enheter levert av forskere når avgrense enheter på tvers av populasjons arter kontinuum. For eksempel anser alle jurisdiksjoner arter anerkjent av taksonomer, som sentrale enheter for bevaring (Tabell 1), men det er fortsatt bred debatt blant taksonomer rundt artsbegreper, og hvordan arter diagnostiseres i praksis varierer innenfor og på tvers av grupper. Dette har ført til en nylig oppfordring til en mer regulert tilnærming til taksonomi (Garnett og Christidis, 2017), som andre har motvirket at regulering av taksonomi ikke er svaret (Thomson et al., 2018). Selv om vi er enige med noen av punktene Gjort Av Thomson et al. (2018) vi foreslår at det er ganske naivt å foreslå at lovgivningen selv må endres for å eksplisitt referere til det spesifikke taxonkonseptet som er underforstått av et navn. Disse og tidligere slike debatter fremhever behovet for evolusjonære biologer og taksonomer å ta ledelsen for å sikre klarhet (åpenhet) i hvordan de oversetter genomisk og annet bevis til avgrensning av arter eller intraspesifikke enheter, og hvordan disse taxa relaterer seg til relevant lovgivning for bevaring og beskyttelse.
Oversettelse Til Ledelsen
som diskutert ovenfor anerkjenner all lovgivning og direktiver artsnivået som den grunnleggende bevaringsenheten, men sørger også for anerkjennelse av ulike infra-spesifikke enheter. Mange har også et underforstått krav til formell navngivning og taksonomisk beskrivelse av arter når de vurderes for bevaringsprioritering og formell beskyttelse. For EKSEMPEL, under Iucn Red Listing Guidelines notering av ikke navngitte arter er motet, og det er et krav om at den nye arten beskrivelse vil bli publisert innen fire år etter at arten blir inkludert på IUCN Rødliste eller det ville bli fjernet. Dette fremhever betydningen av taksonomisk navngivning og beskrivelse av enheter, både på arter og infra-spesifikke nivåer, den begrensede taksonomiske informasjonen for mange plante-og dyregrupper (Mace, 2004; Oliver og Lee, 2010; Wilson, 2017), og behovet for å akselerere taksonomisk innsats, spesielt i biodiversitetsrike områder (Riedel et al., 2013; Wege et al.(2015; Wilson, 2017).Taksonomisk navngitte arter og intraspesifikke taksonomiske kategorier, som underarter og varianter, gir generelt de grunnleggende enhetene for formell notering, bevaringsprioritering og beskyttelse, men i noen jurisdiksjoner er andre kategorier av intraspesifikke bevaringsenheter formelt anerkjent. Disse er generelt identifisert ved bruk av kombinerte genomiske og fenotypiske data. FOR EKSEMPEL anerkjenner Usa Endangered Species Act Distinkt Population Segments (Dps) som er bredt basert På ESUs (Waples, 1991), mens Canadian Species at Risk Act gjenkjenner DUs. En operativ tolkning av «distinkte populasjoner» under Den Kanadiske Arter I Fare Act, DUs tar sikte på å identifisere diskrete sett av populasjoner som representerer viktige komponenter i den evolusjonære arven av arter og som er usannsynlig å bli erstattet gjennom naturlig spredning (Mee et al., 2015).
både DUs og Dps overlapper konseptuelt Med ESUs (Waples et al., 2013) og har blitt brukt mest på nordamerikansk fisk, inkludert nordvest – Usa Laksefisk (Waples, 1995) og Kanadiske coregonids(Mee et al., 2015) Interessant Dps i USA gjelder bare for vertebrater mens I Canada Kan DUs brukes på enhver organisme som kan listes under Sin Handling (se Waples et al., 2013), med DUs for tiden oppført for leddyr, bløtdyr og planter. I Australia, Som Canada, kan forskjellige populasjoner bli oppført for enhver organisme som er anerkjent under Environment Protection and Biodiversity Conservation Act 1999 (EPBC Act), selv om DE ikke er en spesifikt navngitt kategori, for Eksempel DUSs og Dps. Men til dags dato har de bare blitt anerkjent for vertebrater. I USA er det 127 virveldyr Dps oppført for bevaring (https://ecos.fws.gov/ecp/); I Canada Er 70 DUs oppført som dekker alle organismer (http://www.registrelep-sararegistry.gc.ca/species/schedules_e.cfm?id=1); Og I Australia har populasjoner av 7 vertebrate arter som dekker pattedyr, fisk og fugler blitt anerkjent som å ha spesifikk bevaringsstatus under EPBC-Loven (http://www.environment.gov.au/cgi-bin/sprat/public/publicthreatenedlist.pl). Mens vurderinger av disse populasjonene i USA og Canada generelt bruker genetisk informasjon (Waples et al., 2013), I Australia er de vanligvis basert på geografisk og jurisdiksjonell identifikasjon, men i noen av disse tilfellene har de blitt støttet av analyse av genetisk differensiering for pattedyr (Firestone et al ., 1999; Armstrong, 2006) og fish (Colgan og Paxton, 1997; Ahonen et al., 2009).oppføringen av infrarød-spesifikke bevaringsenheter for planter og hvirvelløse dyr i Canada (DUs), men ikke I Usa eller Australia fremhever et annet problem som ikke er spesifikt relatert til avgrensning av bevaringsenheter, men i større grad til betydelige uoverensstemmelser i prioritering av bevaring av biologisk mangfold; ikoniske vertebrater som fugler, pattedyr og fisk er et fokus i mange jurisdiksjoner, mens andre organismer ikke er det. Denne fortrinnsrettsoppføringen av populasjoner av visse organismer gjenspeiler en rekke problemer som er mindre å gjøre med vitenskapen om å avgrense bevaringsenheter og mer å gjøre med oppføring av populasjoner basert på offentlig bevissthet, politisk hensiktsmessighet og den karismatiske naturen til organismen som skal listes (se Waples et al., 2013).Mens I Australia er noen populasjoner av kjente pattedyr, fugler og fisk beskyttet UNDER EPBC-Loven, er det nå en betydelig mengde informasjon basert på molekylære studier i reptiler, hvirvelløse dyr og planter der populasjoner kan gjenkjennes Som ESUs eller MUs, og mange av disse er av bevaringsbetydning gitt deres smale geografiske rekkevidde, liten populasjonsstørrelse, nedgang og følsomhet for truende prosesser. For eksempel kan slående eksempler på høyt strukturerte populasjoner og dypt divergerende linjer innen arter som lett passer til kravet TIL EN ESU, finnes i en rekke lavspredningsvertebrater fra tropene (Figur 2) og andre steder. Tilsvarende i Australske virvelløse dyr, slik som amfipoder og tusenbein, betydelig genetisk struktur, og fylogeografiske mønstre støtte avgrensning av diskrete populasjonsklynger Som Esu som er geografisk ulike, med smale geografiske områder, begrenset til bestemte habitater og representert ved bare noen få populasjoner (Finston et al., 2007; Murphy et al., 2013; Nistelberger et al.( 2014a). Ofte er disse kortdistanse endemiske taxa av bevaringshensyn (Harvey et al., 2011) og er samlokalisert i endemism hotspots (F.eks. Oliver et al., 2017), som selv er høye prioriteringer for multi-taxon bevaring oppmerksomhet (Rosauer Et al., 2018), som Kimberley-Regionen, Og Stirling Range i sørvest Vest-Australia.I Australske planter er det også et betydelig antall populasjonsgenetiske og fylogeografiske studier som identifiserer forskjellige befolkningsgrupper innenfor en rekke arter fra forskjellige slekter som reflekterer disjunct og historisk isolerte befolkningssystemer, geologiske og edafiske kompleksiteter, og opptar kontrasterende habitater når det gjelder vegetasjon og klima (se Broadhurst et al., 2017). Genetisk distinkte populasjoner er spesielt tydelige i arter fra sørvest Vest-Australia (Coates, 2000; Byrne et al., 2014) med en rekke studier som fremhever betydelig genetisk struktur og svært divergerende linjer i en rekke planteslekter og familier, inkludert: Acacia (Mimosaceae; Coates, 1988), Atriplex( Amaranthaceae; Shepherd et al., 2015), Banksia (Proteaceae; Coates et al., 2015), Lambertia (Proteaceae; Byrne et al., 2002), Hakea (Proteaceae; Sampson et al., 2014), Eremophila (Scrophulariaceae; Llorens et al., 2015), Pultanea (Fabaceae); Millar Og Byrne, 2013), Og Calothamnus (Myrtaceae; Nistelberger et al.( 2014b). I mange tilfeller er disse populasjonene anerkjent i bevaringsplanlegging Av Statlige myndigheter og gitt hensyn til miljøkonsekvensvurderinger (se Shepherd et al., 2015) selv om de ikke er oppført for beskyttelse etter lovgivningen. Vi anerkjenner imidlertid at formell anerkjennelse av disse enhetene vil forbedre bevaringen på lang sikt.
Disse Australske eksemplene fremhever det økende behovet for å gjenkjenne dette laget av genetisk mangfold under artsnivået. Utfordringen er å sikre at svært strukturerte populasjoner og dypt divergerende linjer som er av bevaring bekymring er beskyttet i møte med konkurrerende krav om bevaring oppmerksomhet. Utover ikoniske arter er denne utfordringen også tydelig globalt med unntak Av Canada. Unnlatelse av å beskytte klart avgrensede bevaringsenheter vil trolig øke risikoen for kryptisk utryddelse og tap av betydelig genetisk mangfold, som i mange tilfeller kan innebære linjer og historisk isolerte populasjoner som har vedvart innen arter i millioner av år (Moritz, 2002).
Konklusjoner og Anbefalinger
listen over arter og ulike infra-spesifikke kategorier under truet art lovgivning eller under internasjonale organisasjoner som iucn, indikerer en forpliktelse til å beskytte genetisk mangfold på tvers av arter populasjonen kontinuum. Men nåværende tilnærminger til bevaring av biologisk mangfold fokuserer i stor grad på arten slutten av dette kontinuumet med mindre oppmerksomhet gitt til genetisk mangfold og infrarød variasjon. Dette mer begrensede fokuset på genetisk variasjon på intra-artsnivå tyder på at mens bevaringsforvaltere i økende grad anerkjenner verdien av eksplisitt å vurdere genetisk informasjon i ledelseshandlinger og prioritering, mangler de ofte handlingsbar informasjon fra biodiversitetsforskere (Waples et al., 2008; Laikre et al., 2010; Cook og Sgrò, 2017; Taylor et al., 2017). Dette gapet mellom vitenskap og bevaringspraksis hindrer en mer effektiv global tilnærming til å bevare og håndtere genetisk mangfold.Vi foreslår at mangelen på klarhet fra biodiversitetsforskere over bevaringsenheter i stor grad er basert på inkonsekvenser på to nivåer. Den første gjelder flere artsbegreper og de ulike diagnostiske metodene som brukes til å avgrense arter som skaper inkonsekvens på tvers av taksonomiske grupper. Den andre er relatert til mangel på konsistens og avtale av taksonomer, evolusjonære biologer og bevaringsbiologer om hvordan man behandler infra-spesifikke taxa og andre enheter som ESUs, DUs og MUs.
Mens vi anser at lovgivningen ofte allerede er tilstrekkelig fleksibel med hensyn til hvordan taxa (eller «arter») defineres, argumenterer vi for at det er behov for systematisk å adressere populasjonsartkontinuumet, og tydeligere definere enheter for bevaring med større anerkjennelse gitt til begreper SOM ESU og hvordan disse relaterer seg til relevant lovgivning. Vi legger vekt på at dette problemet uten tvil vil bli mer kritisk gitt genomikkens kraft til å oppdage differensiering på gradvis finere skalaer og dens økende bruk for å avgrense arter og infra-spesifikke enheter.
Fra den foregående analysen tilbyr vi følgende anbefalinger:
(i) Genomiske data bør tolkes konservativt når de avgrenser arter, ved hjelp av en integrativ taksonomisk tilnærming som vanligvis inneholder bevis utover de genetiske dataene alene.
(ii) når genetikere identifiserer flere linjer eller distinkte populasjoner innenfor en art, og disse ikke garanterer høyde til artsnivå og ikke er taksonomisk anerkjent, at det er sterkere drivkraft og en mer ensartet tilnærming til å identifisere klart begrunnede intraspesifikke enheter for praktisk vurdering av bevaringsstatus eller konsekvensutredninger, og at disse eksplisitt anerkjennes som enheter for bevaring i sammenheng med lovgivning som er relevant for den jurisdiksjonen.
(iii) Vurdering gis av IUCN, gjennom Sin Conservation Genetics Specialist Group og dens regionale undergrupper, til å utvikle retningslinjer for å gjenkjenne og merking intraspesifikke enheter som samsvarer med beste vitenskapelige praksis.
(iv) den mer nyanserte utsikten som kommer fra evolusjonær genomikk av arter og kontinuumet fra populasjoner til arter, bør kommuniseres mer effektivt av forskere til bevaringsforvaltere og beslutningstakere.
mer generelt ekko vi anbefalinger som ble gjort nylig i en rekke vurderinger (Frankham, 2010; Santamarí og Mé, 2012; Hoban et al., 2013; Moritz Og Potter, 2013; Laity et al., 2015; Pierson et al., 2016; Cook og Sgrò, 2017; Taylor et al., 2017) som fremhever behovet for forskere for å forbedre kommunikasjonen og øke bevisstheten med beslutningstakere, myndigheter, ikke-statlige organisasjoner og utøvere som kommer fra molekylær systematikk, molekylær taksonomi, fylogeografi og befolkningsgenetisk forskning, kan gi kritisk informasjon om bevaring av genetisk mangfold og styring av evolusjonære prosesser.
Forfatterbidrag
DC, MB og CM bidro alle til utarbeidelse og revisjon av manuskriptet.
Interessekonflikt
forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller økonomiske forhold som kan tolkes som en potensiell interessekonflikt.
Takk
dette prosjektet ble støttet Av Australian Research Council (ARC Linkage Grant No. LP120200063), Australian National University og Det Vestlige Australske Departementet For Biologisk Mangfold, Bevaring og Attraksjoner.Afonso Silva, A. C., Bragg, J. G., Potter, S., Fernandes, C., Coelho, M. M., Og Moritz, C. (2017a). Tropisk spesialist versus klima generalist: diversifisering og demografisk historie av søsterarter Av Carlia skinks fra nordvestlige Australia. Mol. Ecol. 26:4045–4058. doi: 10.1111 / mec.14185
CrossRef Full Text/Google Scholar
Afonso Silva, Ac, Santos, N., Ogilvie, H. A., Og Moritz, C. (2017b). Validering og beskrivelse av to nye nordvestlige Australske Rainbow skinks med multispecies coalescent metoder og morfologi. PeerJ 5: e3724. doi: 10.7717 / peerj.3724
PubMed Abstrakt / CrossRef Fulltekst/Google Scholar
Ahonen, H., Harcourt, R. G., Og Stow, A. J. (2009). Nuclear OG mitokondrielt DNA avslører isolasjon av imperilled grå sykepleier hai populasjoner (Carcharias taurus). Mol. Ecol. 18, 4409–4421. doi: 10.1111 / j.1365-294X. 2009. 04377.x
PubMed Abstract / CrossRef Full Text/Google Scholar
Armstrong, K. n. (2006). Phylogeographic struktur I Rhinonicteris aurantia( Chiroptera: Hipposideridae): implikasjoner for bevaring. Acta Chiropterologica 8, 63-81. doi: 10.3161/1733-5329(2006)82.0.CO
CrossRef Full Text | Google Scholar
Arnold, M. L., Og Kunte, K. (2017). Adaptiv genetisk utveksling: en sammenflettet historie av blanding og evolusjonær innovasjon. Trender Ecol. Evol. 32, 601–611. doi: 10.1016 / j.tre.2017.05.007
PubMed Abstract / CrossRef Full Text / Google Scholar
Avise, J. c. (2004). Molekylære Markører, Naturhistorie og Evolusjon. 2. Edn. Sunderland, MA; Oxford: Oxford University Press.Avise, J. C., Arnold, J., Ball, R. M., Bermingham, E., Lamb, T., Neigel, J. E., Et al. (1987). Intraspesifikk fylogeografi: mitokondrielt DNA-bro mellom populasjonsgenetikk og systematikk. Anne. Rev. Ecol. Sys. 18, 489–522. doi: 10.1146/annurev.es.18.110187.002421
CrossRef Full Text | Google Scholar
Bickford, D., Lohman, D. J., Sodhi, N. S, Ng, P. K., Meier, R., Winker, K., Et al. (2007). Kryptiske arter som et vindu på mangfold og bevaring. Trender Ecol. Evol. 22, 148–155. doi: 10.1016 / j.tre.2006.11.004
PubMed Abstract | CrossRef Full Text/Google Scholar
Blom, M. P. K., Bragg, J. G., Potter, S., Og Moritz, C. (2016). Regnskap for usikkerhet i gene tree estimering: sammendrag-coalescent arter tre slutning i en utfordrende stråling Av Australske øgler. Syst. Biol. 66, 352–366. doi: 10.1093/sysbio/syw089
PubMed Abstract | CrossRef Full Text | Google Scholar
Broadhurst, L., Breed, M., Lowe, A., Bragg, J., Catullo, R., Coates, Dj, Et al. (2017). Genetisk mangfold og struktur Av Den Australske floraen. Dykkere. Distribuer. 23, 41–52. doi: 10.1111 / ddi.12505
CrossRef Full Text/Google Scholar
Byrne, M., Coates, D. J., Forest, F., Hopper, S. D., Krauss, S. L., Sniderman, J. M. K., et al. (2014). «En mangfoldig flora – art og genetiske relasjoner,» I Plant Life on The Sandplains I Southwest Australia, Et Globalt Biologisk Mangfold Hotspot, redaktører H. Lambers (Crawley, WA: University Of Western Australia Publishing), 81-99.
PubMed Abstract
Byrne, M., Macdonald, B., Og Coates, Dj (2002). Phylogeographic mønstre og høy cpDNA mangfold innen Acacia acuminata (Leguminosae:Mimosoideae). kompleks I Vest-Australia. J. Evol. Biol. 15, 576–587. doi: 10.1046/j.1420-9101. 2002. 00429.X
CrossRef Full Text | Google Scholar
Carstens, B. C., Pelletier, T. A., Reid, N. M., Og Satler, J. D. (2013). Hvordan mislykkes på arter avgrensning. Mol. Ecol. 22, 4369–4383. doi: 10.1111 / mec.12413
PubMed Abstract / CrossRef Full Text / Google Scholar
Coates, Dj (1988). Genetisk mangfold og populasjonsgenstruktur i den sjeldne Chittering grass wattle, Acacia anomala Court. Aust. J. Bot. 36:273–286. doi: 10.1071/BT9880273
Kryssref Fulltekst | Google Scholar
Coates, Dj (2000). Definere bevaringsenheter i en rik og fragmentert flora: implikasjoner for forvaltning av genetiske ressurser og evolusjonære prosesser i sør-vest Australia planter. Aust. J. Bot. 48, 329–339. doi: 10.1071/BT99018
CrossRef Full Text | Google Scholar
Coates, Dj, McArthur, Sl, Og Byrne, M. (2015). Betydelig genetisk mangfold tap etter patogen drevet populasjons utryddelse i den sjeldne endemiske Banksia brownii (Proteaceae). Biol. Conserv. 192, 353–360. doi: 10.1016 / j.biocon.2015.10.013
CrossRef Full Text / Google Scholar
Colgan, Dj, Og Paxton, J. R. (1997). Biokjemisk genetikk og anerkjennelse av en vestlig bestand av felles gemfish, Rexea solandri (Scombroidea: Gempylidae)., I Australia. Mar. Freshw. Res. 48, 103-118. doi: 10.1071 / MF96048
CrossRef Full Text / Google Scholar
Konvensjonen Om Biologisk Mangfold (2007). Tilgjengelig online på:https://www.cbd.int/doc/meetings/cop-bureau/cop-bur-2007/cop-bur-2007-10-14-en.pdf
Cook, C. N., og Sgrò, C. M. (2017). Samkjøre vitenskap og politikk for å oppnå evolusjonært opplyst bevaring. Conserv. Biol. 31, 501–512. doi: 10.1111 / cobi.12863
PubMed Abstract | CrossRef Full Text/Google Scholar
Cook, L. G., Edwards, Rd, Crisp, Md, Og Hardy, N. B. (2010). Trenger morfologi alltid være nødvendig for nye arter beskrivelser? Inverter. Sys. 24, 322–332. doi: 10.1071/IS10011
CrossRef Full Text | Google Scholar
Cracraft, J. (1983). Artsbegreper og artsanalyse. Curr. Ornithol. 1, 159–187. doi: 10.1007/978-1-4615-6781-3_6
CrossRef Full Text | Google Scholar
Crandall, K. A., Bininda-Emonds, O. R., Mace, G. M. og Wayne, R. K. (2000). Vurderer evolusjonære prosesser i bevaringsbiologi. Trender Ecol. Evol. 15, 290–295. doi: 10.1016 / S0169-5347 (00)01876-0
PubMed Abstract / CrossRef Full Text / Google Scholar
Kutter, A. D., Og Payseur, Ba (2013). Genomiske signaturer av utvalg på koblede områder: samlende ulikhet mellom arter. Nat. Pastor Genet. 14, 262–274. doi: 10.1038 / nrg3425
PubMed Abstrakt / CrossRef Full Tekst / Google Scholar
Dayrat, B. (2005). Integrativ taksonomi. Biol. J. Linn. Soc. 85, 407–415. doi: 10.1111/j.1095-8312.2005. 00503.x5
CrossRef Full Text/Google Scholar
de Queiroz, K. (1998). «Den generelle avstamning begrepet arter, arter kriterier, og prosessen med artsdannelse,» I Former: Arter og Artsdannelse, D. J. Howard, R. H. Berlocher (New York, NY: Oxford University Press), 57-75.
Google Scholar
de Queiroz, K. (2007). Artsbegreper og artsavgrensning. Syst. Biol. 56, 879–886. doi: 10.1080/10635150701701083
PubMed Abstract / CrossRef Full Text / Google Scholar
Degnan, J. H., Og Rosenberg, N. A. (2009). Gent tree discordance, fylogenetisk slutning og multispecies coalescent. Trender Ecol. Evol. 24, 332–340. doi: 10.1016 / j.tre.2009.01.009
PubMed Abstract | CrossRef Full Text/Google Scholar
Devey, D. S., Bateman, R. M., Fay, M. F., Og Hawkins, J. A. (2008). Venner eller slektninger? Fylogenetikk og arter avgrensning i den kontroversielle Europeiske orkide slekten Ophrys. Anne. Robot. 101, 385–402. doi: 10.1093/aob/mcm299
PubMed Abstract | CrossRef Full Text | Google Scholar
Dizon, A. E., Lockyer, C, Perrin, W. F., Demaster, D. P. Og Sisson, J. (1992). Rethinking lager konseptet – en fylogeografisk tilnærming. Conserv. Biol. 6, 24–36. doi: 10.1046/j.1523-1739. 1992. 610024.X
CrossRef Full Text/Google Scholar
Doughty, P., Bourke, G., Tedeschi, L., Pratt, R., Oliver, P., Palmer, R., Et al. (2018). Arten er avgrenset i Gehyra nana (Squamata: Gekkonidae). kompleks: kryptisk og divergerende morfologisk utvikling i De Australske Monsoonale Tropene, med beskrivelsen av fire nye arter. Zootaxa, 4403: 201-244. doi: 10.11646 / zootaxa.4403.2.1
PubMed Abstract | CrossRef Full Text/Google Scholar
Dynesius, M., Og Jansson, R. (2014). Persistens Av arter i arter: en forsømt kontroll av artsfrekvenser. Evolusjon 68, 923-934. doi: 10.1111 / evo.12316
PubMed Abstrakt / CrossRef Fulltekst/Google Scholar
Edwards, S. V., Potter, S., Schmitt, C. J., Bragg, J. G., Og Moritz, C. (2016). Retikulering, divergens og fylogeografi–fylogenetikkkontinuumc. Proc. Natl. Acad. Sci. U. S. A. 113, 8025-8032. doi: 10.1073 / pnas.1601066113
CrossRef Full Tekst / Google Scholar
Tro, D. P. (1992). Bevaringsevaluering og fylogenetisk mangfold. Biol. Svindel. 61, 1–10. doi: 10.1016/0006-3207(92)91201-3
CrossRef Full Text | Google Scholar
Finston, T. L., Johnson, Ms, Humphreys, Wf, Eberhard, S. M., Og Halse, S. A. (2007). Kryptisk artsdannelse i to utbredte underjordiske amphipod slekter reflekterer historiske drenering mønstre i et gammelt landskap. Mol. Ecol. 16, 355–365. doi: 10.1111 / j.1365-294X. 2006. 03123.X
PubMed Abstract | CrossRef Full Text | Google Scholar
Firestone, K. B., Elphinstone, Ms, Sherwin, W. B., Og Houlden, B. a. (1999). Tigerkulene Dasyurus maculatus (Dasyuridae: Marsupalia) er en rovfugl som er i fare. Mol. Ecol. 8, 1613–1625. doi: 10.1046/j.1365-294x.1999. 00745.X
CrossRef Full Text | Google Scholar
Fiš, C., Robinson, Ct, Og Malard, F. (2018). Kryptiske arter som et vindu inn i paradigmeskiftet av artsbegrepet. Mol. Ecol. 27, 613–635. doi: 10.1111 / mec.14486
PubMed Abstrakt | CrossRef Fulltekst/Google Scholar
Frankham, R. (2010). Utfordringer og muligheter for genetiske tilnærminger til biologisk bevaring. Biol. Conserv. 143,1919–1927. doi: 10.1016 / j.biocon.2010.05.011
CrossRef Full Text/Google Scholar
Frankham, R., Ballou, J. D., Dudash, M. R., Eldridge, M. D., Fenster, C. B., Lacy, R. C., et al. (2012). Implikasjoner av ulike artsbegreper for å bevare biologisk mangfold. Biol. Conserv. 153, 25–31. doi: 10.1016 / j.biocon.2012.04.034
CrossRef Full Text/Google Scholar
Fraser, Dj, Og Bernatchez, L. (2001). Adaptiv evolusjonær bevaring: mot et enhetlig konsept for å definere bevaringsenheter. Mol. Ecol. 10, 2741–2752. doi: 10.1046 / j.1365-294X. 2001.t01-1-01411.X
PubMed Abstract | CrossRef Full Text | Google Scholar
Funk, W. C., Mckay, J. K., Hohenlohe, P. A., Og Allendorf, F. W. (2012). Utnytte genomikk for avgrense bevaring enheter. Trender Ecol. Evol. 27, 489–496. doi: 10.1016 / j.tre.2012.05.012
PubMed Abstract | CrossRef Full Text/Google Scholar
Garnett, S. T., Og Christidis, L. (2017). Taksonomi anarki hemmer bevaring. Natur 546, 25-27. doi: 10.1038/546025a
PubMed Abstract | CrossRef Full Text | Google Scholar
Gippoliti, S., Cotterill, Fpd, Zinner, D., Og Groves, Cp (2018). Virkninger av taksonomisk treghet for bevaring Av Afrikansk ungulatmangfold: oversikt. Biol. Åp 93, 115-130. doi: 10.1111 / brv.12335
PubMed Abstract | CrossRef Full Text/Google Scholar
Grant, V. (1981). Plante Spesiering. New York: Columbia University Press.s.
PubMed Abstract / Google Scholar
Hamilton, C. W., Og Reichard, S. H. (1992). Nåværende praksis i bruk av underarter, varianter og forma i klassifisering av ville planter. Taxon 41, 485-498. doi: 10.2307/1222819
CrossRef Full Text | Google Scholar
Harvey, M. S., Rix, M. G., Framenau, V. W., Hamilton, Z. R., Johnson, M. S., Teale, R. J., et al. (2011). Beskytte uskyldige: studere kort rekkevidde endemisk taxa forbedrer bevaring utfall. Inverter. Syst. 25, 1–10. doi: 10.1071/IS11011
CrossRef Full Text | Google Scholar
Hei, J., Waples, Rs, Arnold, Ml, Butlin, Rk, Og Harrison, Rg (2003). Forstå og konfrontere arter usikkerhet i biologi og bevaring. Trender Ecol. Evol. 18, 597–603. doi: 10.1016 / j.tre.2003.08.014
CrossRef Full Text/Google Scholar
Hoban, S. M., Hauffe, H. C., Perez-Espona, S., Arntzen, J. W., Bertorelle, G., Bryja, J., et al. (2013). Bringe genetisk mangfold i forkant av bevaring politikk og forvaltning. Conserv. Genet. Resour. 5, 593–598. doi: 10.1007 / s12686-013-9859-y
CrossRef Full Text | Google Scholar
Isaac, Nj, Mallet, J., og Mace, Gm (2004). Taksonomisk inflasjon: dens innflytelse på makroøkologi og bevaring. Trender Ecol. Evol. 19, 464–469. doi: 10.1016 / j.tre.2004.06.004
PubMed Abstract / CrossRef Full Text / Google Scholar
Isaac, N. J., Turvey, S. T., C., Waterman, C., Og Baillie, J. E. (2007). Pattedyr på KANTEN: bevaringsprioriteter basert på trussel og fylogeni. PLoS ONE 2: e296. doi: 10.1371 / tidsskrift.pone.0000296
PubMed Abstract | CrossRef Full Text/Google Scholar
Jones, Mr, Og God, Jm (2016). Målrettet fangst i evolusjonær og økologisk genomikk. Mol. Ecol. 25, 185–202. doi: 10.1111 / mec.13304
PubMed Abstract | CrossRef Full Text/Google Scholar
Jorger, K. M., Og Schrö, M. (2013). Hvordan beskrive en kryptisk art? Praktiske utfordringer i molekylær taksonomi. Front. Zool. 10:59. doi: 10.1186/1742-9994-10-59
PubMed Abstract | CrossRef Full Text/Google Scholar
Laikre, L., Allendorf, F., Aroner, L. C., Baker, S., Gregovich, D. P., Hansen, M. M., Et al. (2010). Forsømmelse av genetisk mangfold i gjennomføringen av konvensjonen om biologisk mangfold. Conserv. Biol. 24, 86–88. doi: 10.1111/j.1523-1739.2009. 01425.X
PubMed Abstract / CrossRef Full Text / Google Scholar
Laikre, L., Palm, S., Og Ryman, N. (2005). Genetisk populasjonsstruktur av fisk: implikasjoner for kystsoneforvaltning. Ambio 34111-119. doi: 10.1579/0044-7447-34.2.111
PubMed Abstract | CrossRef Full Text/Google Scholar
Laity, T., Laffan, S. W., Gonz@lez-Orozco, C. E., Faith, D. P., Rosauer, D. F., Byrne, M., et al. (2015). Phylodiversity å informere bevaring politikk: En Australsk eksempel. Sci. Totalt Miljø. 534, 131–143. doi: 10.1016 / j.scitotenv.2015.04.113
PubMed Abstract / CrossRef Full Text / Google Scholar
Leache, A. D., Og Oaks, J. R. (2017). Nytten av enkelt nukleotidpolymorfisme (SNP). data i fylogenetikk. Anne. Rev. Ecol. Syst. 48, 69–84. doi: 10.1146/annurev-ecolsys-110316-022645
CrossRef Full Text | Google Scholar
Llorens, T. M., Macdonald, B., McArthur, S., Coates, D. J., and Byrne, M. (2015). Disjunct, highly divergent genetic lineages within two rare Eremophila (Scrophulariaceae: Myoporeae). species in a biodiversity hotspot: implications for taxonomy and conservation. Biol. J. Linn. Soc. 177, 96–111. doi: 10.1111/boj.12228
CrossRef Full Text | Google Scholar
Love Stowell, S. M., Pinzone, C. A., and Martin, A. P. (2017). Overvinne barrierer for aktive tiltak for genetisk mangfold. Biodrivere. Conserv. 26, 1753–1765. doi: 10.1007 / s10531-017-1330-z
CrossRef Full Text/Google Scholar
Mace, G. M. (2004). Taksonomiens rolle i bevaring av arter. Philos. Trans. R. Soc. B: Biol. Sci. 359, 711–719. doi: 10.1098 / rstb.2003.1454
PubMed Abstract / CrossRef Full Text / Google Scholar
Mallet, J. (2007). Hybrid spesiering. Natur 446, 279-283. doi: 10.1038/nature05706
PubMed Abstrakt | CrossRef Full Tekst/Google Scholar
Mayr, E. (1942). Systematikk og Artenes Opprinnelse. New York: Columbia University Press.s.Google Scholar
Mee, J. A., Bernatchez, L., Reis, J. D., Rogers, S. M. og Taylor, E. B. (2015). Identifisere betegnbare enheter for intraspesifikk bevaringsprioritering: en hierarkisk tilnærming anvendt på lake sik artskomplekset (Coregonus spp.). Evol. Appl. 8, 423–441. doi: 10.1111 / eva.12247
PubMed Abstract | CrossRef Full Text | Google Scholar
Millar, M. A., and Byrne, M. (2013). Cryptic divergent lineages of Pultenaea pauciflora M.B. Scott (Fabaceae: Mirbelieae) exhibit different evolutionary history. Biol. J. Linn. Soc. 108, 871–881. doi: 10.1111/bij.12002
CrossRef Full Text | Google Scholar
Moritz, C. (1994). Defining evolutionarily significant units for conservation. Trends Ecol. Evol. 9, 373–375. doi: 10.1016/0169-5347(94)90057-4
CrossRef Full Text | PubMed Abstract
Moritz, C. (1999). Bevaringsenheter og translokasjoner: strategier for bevaring av evolusjonære prosesser. Hereditas 130, 217-228. doi: 10.1111/j.1601-5223. 1999. 00217.x
CrossRef Full Text / Google Scholar
Moritz, C. (2002). Strategier for å beskytte biologisk mangfold og de evolusjonære prosessene som opprettholder det. Syst. Biol. 51, 238–254. doi: 10.1080/10635150252899752
PubMed Abstract / CrossRef Full Text / Google Scholar
Moritz, C., Og Potter, S. (2013). Betydningen av et evolusjonært perspektiv i bevaringspolitisk planlegging. Mol. Ecol. 22, 5969–5971. doi: 10.1111 / mec.12565
PubMed Abstract | CrossRef Full Text/Google Scholar
Moritz, C., Pratt, R. C., Bank, S., Bourke, G., Bragg, J. G., Doughty, P., et al. (2018). Kryptisk avstamning mangfold, kroppsstørrelse divergens, og sympatry i en art kompleks Av Australske øgler (Gehyra). Evolusjon 27, 54-66. doi: 10.1111 / evo.13380
CrossRef Full Text/Google Scholar
Morrison, W. R., Lohr, J. L., Duchen, P., Wilches, R., Trujillo, D., Mair, M., et al. (2009). Virkningen av taksonomisk endring på bevaring: Dreper det, kan det redde, eller er det bare irrelevant? Biol. Svindel. 142, 3201–3206. doi: 10.1016 / j.biocon.2009.07.019
CrossRef Full Text/Google Scholar
Murphy, N. P., Adams, M., Guzik, Mt, Og Austin, A. D. (2013). Ekstraordinær mikro-endemisme I Australske ørkenen våren amphipods. Mol. Fylogenet. Evol. 66, 645–653. doi: 10.1016 / j. ympev.2012.10.013
PubMed Abstract / CrossRef Full Text / Google Scholar
Murphy, N. P., King, R. A. og Delean, S. (2015). Arter, ESUs eller populasjoner? Avgrense og beskrive morfologisk kryptisk mangfold I Australske ørkenen våren amphipods. Inverter. Syst. 29, 457–467. doi: 10.1071/IS14036
CrossRef Full Text | Google Scholar
Nater, A., Mattle-Greminger, M. P., Nurcahyo, A., Nowak, Mg, De Manual, M., Desai, T., Et al. (2017). Morfometrisk, atferdsmessig og genomisk bevis for en ny orangutangart. Curr. Celle Biol. 27, 3487–3498. doi: 10.1016 / j.cub.2017.09.047
CrossRef Full Text/Google Scholar
Nicotra, A. B., Chong, C., Bragg, J. G., Ong, C. R., Aitken, N. C., Chuah, A., et al. (2016). Populasjon og fylogenomisk dekomponering via genotyping-by-sekvensering I Australsk Pelargonium. Mol. Ecol. 25, 2000–2014. doi: 10.1111 / mec.13584
PubMed Abstract | CrossRef Full Text/Google Scholar
Nistelberger, H., Byrne, M., Coates, D. ,Og Roberts, J. D. (2014a). Sterk fylogeografisk struktur i en tusenbein indikerer Pleistocen vikarians mellom populasjoner på banded jernformasjoner i halvtørre Australia. PLoS EN 9: e0093038. doi: 10.1371 / tidsskrift.pone.0093038
PubMed Abstract | CrossRef Full Text/Google Scholar
Nistelberger, H., Gibson, N., Macdonald, B., Tapper, S.-L., Og Byrne, M. (2014b). Phylogeographic bevis for to mesic refugia i et biologisk mangfold hotspot. Arvelighet 113, 454-463. doi: 10.1038 / hdy.2014.46
PubMed Abstract | CrossRef Full Text/Google Scholar
Ogilvie, H. A., Heled, J., Xie, D., Og Drummond, A. J. (2016). Computational ytelse og statistisk nøyaktighet av beast og sammenligninger med andre metoder. Syst. Biol. 65, 381–396. doi: 10.1093 / sysbio / syv118
PubMed Abstract / CrossRef Full Text / Google Scholar
Oliver, Pm, Keogh, Js, Og Moritx, C. (2015). Nye tilnærminger til katalogisering og forståelse av evolusjonært mangfold: et perspektiv fra Australsk herpetologi. Aust. J. Zool. 62, 417–430. doi: 10.1071/ZO14091
CrossRef Full Text | Google Scholar
Oliver, Pm, Laver, Rj, Demello Martins, F., Pratt, Rc, Hunjan, S. og Moritz, C. (2017). En roman hotspot av vertebrate endemisme og et evolusjonært refugium i tropisk Australia. Dykker. Distribuer. 23, 53–66. doi: 10.1111 / ddi.12506
CrossRef Full Text/Google Scholar
Oliver, Pm, Og Lee, Msy (2010). De botaniske og zoologiske kodene hindrer biodiversitetsforskning ved å motvirke publisering av ikke navngitte nye arter. Skatt 59, 1201-1205. doi: 10.2307 / 20773990
CrossRef Full Text/Google Scholar
Padial, J., Miralles, A., De La Riva, I., Og Vences, M. (2010). Den integrerende fremtiden for taksonomi. Front. Zool. 7:16. doi: 10.1186/1742-9994-7-16
PubMed Abstract | CrossRef Full Text/Google Scholar
Palsb@ll, P. J., Bé Rubé, M., Og Allendorf, F. W. (2007). Identifisering av forvaltningsenheter ved hjelp av populasjonsgenetiske data. Trender Ecol. Evol. 22, 11–16. doi: 10.1016 / j.tre.2006.09.003
PubMed Abstract / CrossRef Full Text/Google Scholar
Patton, J. L., Og Smith, M. F. (1994). Paraphyly, polyphyly, og arten av arter grenser I Lomme Gophers (Slekten Thomomys). Syst. Biol. 43, 11–26. doi: 10.1093 / sysbio / 43.1.11
CrossRef Full Text/Google Scholar
Pierson, J. C., Coates, D. J., Oostermeijer, J. G. B., Beissinger, S. R., Bragg, J. G., Sunnucks, P., et al. (2016). Genetiske faktorer i truede arter utvinning planer på tre kontinenter. Front. Ecol. Miljø. 14, 433–440. doi: 10.1002 / avgift.1323
CrossRef Full Text/Google Scholar
Potter, S., Bragg, J. G., Peter, Bm, Bi, K., Og Moritz, C. (2016). Fylogenomikk på tips: inferring linjer og deres demografiske historie i en tropisk øgle, Carlia amax. Mol. Ecol. 25, 1367–1380. doi: 10.1111 / mec.13546
PubMed Abstract | CrossRef Full Text/Google Scholar
Potter, S., Xue, At, Bragg, Jg, Rosauer, Df, Roycroft, Ej og Moritz, C. (2018). Pleistocen klimaendringer driver diversifisering over en tropisk savanne. Mol. Ecol. 27, 520–532. doi: 10.1111 / mec.14441/full
PubMed Abstract | CrossRef Full Text/Google Scholar
Rabosky, D. L., Hutchinson, M. N., Donnellan, Sc, Talaba, Al, Og Lovette, I. J. (2014). Fylogenetisk demontering av artsgrenser i en utbredt gruppe Australske skinker (Scincidae: Ctenotus). Mol. Fylogenet. Evol. 77, 71–82. doi: 10.1016 / j. ympev.2014.03.026
PubMed Abstract | CrossRef Full Text/Google Scholar
Ralls, K., Ballou, J. D., Dudash, M. R., Eldridge, M. D. B., Fenster, C. B., Lacy, R. C., et al. (2018). Kall for et paradigmeskifte i genetisk forvaltning av fragmenterte populasjoner. Conserv. Lett. 11, 1–6. doi: 10.1111 / conl.124126
Kryssref Fulltekst / Google Scholar
Rannala, B. (2015). Kunsten og vitenskapen om artsavgrensning. Curr. Zool. 61, 846–853. doi: 10.1093/czoolo/61.5.846
CrossRef Full Text | Google Scholar
Riedel, A., Sagata, K., Suhardjono, Y. R., Tä, R. og Balke, M. (2013). Integrativ taksonomi på fast track-mot mer bærekraft i biodiversitetsforskning. Front. Zool. 10:15. doi: 10.1186/1742-9994-10-15
PubMed Abstract / CrossRef Full Text / Google Scholar
Rieseberg, L. H., And Willis, J. H. (2007). Plante spesiering. Vitenskap 317, 910-914. doi: 10.1126 / vitenskap.1137729
PubMed Abstract | CrossRef Full Text/Google Scholar
Rosauer, D. F., Blom, M. P. K., Bourke, G., Catalano, S., Donnellan, S., Mulder, E., Et al. (2016). Fylogeografi, hotspots og bevaringsprioriteter: et eksempel Fra Den Øverste Enden Av Australia. Biol. Svindel. 204, 83–93. doi: 10.1016 / j.biocon.2016.002
CrossRef Full Text/Google Scholar
Rosauer, D. F., Byrne, M., Blom, M. P. K., Coates, D. J., Donnellan, S., Doughty, P., et al. (2018). Real-world bevaring planlegging for evolusjonært mangfold I Kimberley, Australia, sidesteps usikker taksonomi Conserv. Lett. 11: e12438. doi: 10.1111 / conl.12438
CrossRef Full Text/Google Scholar
Rosenblum, E. B., Sarver, B. A. J., Brown, J. W., Des Roches, S., Hardwick, K. M., Hether, T. D., et al. (2012). Goldilocks møter Santa Rosalia: en flyktig spesiasjonsmodell forklarer mønstre av diversifisering på tvers av tidsskalaer. Evol. Biol. 39, 255–261. doi: 10.1007 / s11692-012-9171-X
PubMed Abstract | CrossRef Full Text | Google Scholar
Roux, C., Fraï, C., Romiguier, J., Anciaux, Y., Galtier, N. og Bierne, N. (2016). Shedding lys på den grå sonen av artsdannelse langs et kontinuum av genomisk divergens. PLoS Biol. 14: e2000234. doi: 10.1371 / tidsskrift.pbio.2000234
PubMed Abstract | CrossRef Full Text/Google Scholar
Ryder, O. A. (1986). Arter bevaring og systematikk-dilemmaet av underarter. Trender Ecol. Evol. 1, 9–10. doi: 10.1016/0169-5347 (86)90059-5
CrossRef Full Text | Google Scholar
Sampson, J. F., Byrne, M., Yates, C. J., Gibson, N., Thavornkanlapachai, R., Stankowski, S., et al. (2014). Moderne pollen-mediert geninnvandring gjenspeiler den historiske isolasjonen av en sjelden busk i et fragmentert landskap. Arvelighet 112, 172-181. doi: 10.1038 / hdy.2013.89
PubMed Abstract | CrossRef Full Text/Google Scholar
Santamarí, L., og Mé, P. F. (2012). Evolusjon i biodiversitetspolitikken: nåværende hull og fremtidige behov. Evol. Appl. 5, 202–218. doi: 10.1111 / j.1752-4571. 2011. 00229.X
PubMed Abstract | CrossRef Full Text | Google Scholar
Schlick-Steiner, B. C., Steiner, F. M., Seifert, B., Stauffer, C., Christian, E., Og Crozier, R. H. (2010). Integrativ taksonomi: en multisource tilnærming til å utforske biologisk mangfold. Annu. Rev. Entomol. 55, 421–438. doi: 10.1146 / annurev-ento-112408-085432
PubMed Abstract | CrossRef Full Text/Google Scholar
Seehausen, O, Takimoto, G., Roy, D., Og Jokela, J. (2008). Artsdannelse reversering og biodiversitet dynamikk med hybridisering i skiftende miljøer. Mol Ecol. 17, 30–44. doi: 10.1111 / j.1365-294X. 2007. 03529.X
PubMed Abstract | CrossRef Full Text | Google Scholar
Shepherd, K. A., Thiele, K. R., Sampson, J., Coates, Dj, Og Byrne, M. (2015). En sjelden, ny Art Av Atriplex (Chenopodiaceae) som består av to genetisk distinkte, men morfologisk kryptiske populasjoner i tørre Vest-Australia: implikasjoner for taksonomi og bevaring. Aust. Syst. Robot. 28, 234–245. doi: 10.1071 / SB15029
CrossRef Fulltekst / Google Scholar
Singhal, S., Og Bi, K. (2017). Historien rydder opp søl: virkningen av tid i driving divergens og introgression i en tropisk sutur sone. Evolusjon 71, 1888-1899. doi: 10.1111 / evo.13278
PubMed Abstract | CrossRef Full Text/Google Scholar
Singhal, S., Hoskin, C., Couper, P., Potter, S., Og Moritz, C. (2018). Rammeverk for å løse kryptiske arter: en casestudie fra øgler Av De Australske våte tropene. Syst. Biol. . doi: 10.1093 / sysbio/syy
PubMed Abstract / CrossRef Full Text / Google Scholar
Singhal, S., Og Moritz, C. (2013). Reproduktiv isolasjon mellom fylogeografiske linjeskalaer med divergens. Proc. Roy. Soc B 280:20132246. doi: 10.1098 / rspb.2013.2246026
PubMed Abstract | CrossRef Full Text/Google Scholar
Steane, D. A., Potts, B. M., McLean, E., Collins, L., Prober, S. M., Lager, W. D., Et al. (2015). Genom-wide skanner avsløre kryptisk populasjonsstruktur i en tørr tilpasset eucalypt. Treet Genet. Genom 11:33. doi: 10.1007 / s11295-015-0864-z
CrossRef Full Text | Google Scholar
Struck, T. H., Feder, J. L., Bendiksby, M., Birkeland, S., Cerca, J., Gusarov, V. I., et al. (2017). Finne evolusjonære prosesser skjult i kryptiske arter. Trender Ecol. Evol. 33, 153–163. doi: 10.1016 / j.tre.2017.11.007
PubMed Abstract / CrossRef Full Text / Google Scholar
Sukumaran, J., Og Knowles, L. l. (2017). Multispecies coalescent avgrenser struktur ikke arter. Proc. Nat. Aca. Sci. U. S. A. 114, 1607-1612. doi: 10.1073 / pnas.1607921114
PubMed Abstract | CrossRef Full Text/Google Scholar
Swarts, Nd, Clements, M. A., Bower, C. C., Og Miller, J. T. (2014). Definere bevaring enheter i et kompleks av morfologisk lignende, seksuelt villedende, svært truede orkideer. Biol. Svindel. 174, 55–64. doi: 10.1016 / j.biocon.2014.03.017
CrossRef Full Text/Google Scholar
Tautz, D., Arctander, P., Minelli, A., Thomas, Rh ,Og Vogler, A. P. (2003). En bønn FOR DNA taksonomi. Trender Ecol. Evol. 18, 70–74. doi: 10.1016 / S0169-5347 (02)00041-1
CrossRef Full Text/Google Scholar
Taylor, H. R., Dussex, N., og van Heezik, Y. (2017). Bygge bro over bevaringsgenetikkgapet ved å identifisere barrierer for implementering for bevaringsutøvere. Glob. Ecol. Conserv. 10, 231–242. doi: 10.1016 / j.gecco.2017.04.001
CrossRef Full Text/Google Scholar
Thomson, S. A., Pyle, R. L., Ahyong, S. T., Alonso-Zarazaga, M., Ammirati, J., Araya, J. F., Et al. (2018). Taksonomi basert på vitenskap er nødvendig for global bevaring. PLoS Biol. 16: e2005075. doi: 10.1371 / journal.pbio.2005075
PubMed Abstract | CrossRef Full Text/Google Scholar
Vieites, D. R., Wollenberg, K. C., Andreonec, F., Kö, J., Glaw, F., Og Vences, M. (2009). Enorme undervurdering Av Madagaskars biologiske mangfold dokumentert av en integrerende amfibie inventar. Proc. Natl. Acad. Sci. U. S. A. 106, 8267-8272. doi: 10.1073 / pnas.0810821106
CrossRef Fulltekst
Vogler, A. P., og de Salle, R. (1994). Diagnostisering Enheter Av Bevaring Ledelse. Svindel. Biol. 8, 354–363. doi: 10.1046/j.1523-1739. 1994. 08020354.x
CrossRef Full Text/Google Scholar
Waples, R. S. (1991). Stillehavslaks, Oncorhynchus spp. og definisjonen av» arter » under Endangered Species Act. Mar. Fisk. Åp 53, 11-22.
Google Scholar
Waples, R. S. (1995). «Evolutionarily significant units and the conservation of Biological diversity under Endangered Species Act,» I Evolusjon og Akvatisk Økosystem: Definere Unike Enheter I Population Conservation, eds Jl Nielsen (Bethesda, MD:American Fisheries Society), 8-27.
Google Scholar
Waples, R. S., Gustafson, R. G., Weitkamp, L. A., Myers, J. M., Jjonson, O. W., Busby, P. J., Et al. (2001). Karakteriserer mangfold i laks Fra Stillehavet Nordvest. J. Fisk Biol. 59, 1–41. doi: 10.1111 / j.1095-8649. 2001.tb01376.X
CrossRef Full Text | Google Scholar
Waples, R. S., Nammack, M., Cochrane, J. F., Og Hutchings, J. A. (2013). En fortelling om to handlinger: truede arter notering praksis I Canada og Usa. Biovitenskap 63, 723-734. doi: 10.1093/bioscience / 63.9.723
CrossRef Full Text | Google Scholar
Waples, Rs, Punt, Ae, Og Cope, Jm (2008). Integrering av genetiske data i forvaltning av marine ressurser: hvordan kan vi gjøre det bedre? Fisk 9, 423-449. doi: 10.1111/j.1467-2979. 2008. 00303.X
CrossRef Full Text | Google Scholar
Wege, J. A., Thiele, K. R., Shepherd, K. A., Butcher, R., Macfarlane, T. D., Og Coates, Dj (2015). Strategisk taksonomi i et biodiverse landskap: en ny tilnærming til å maksimere bevaringsutfall for sjeldne og dårlig kjente flora. Biodrivere. Conserv. 24, 17–32. doi: 10.1007 / s10531-014-0785-4
CrossRef Full Text/Google Scholar
Whitlock, R. (2014). Forholdet mellom adaptivt og nøytralt genetisk mangfold og økologisk struktur og funksjon: en meta-analyse. J. Ecol. 102, 857–872. doi: 10.1111 / 1365-2745.12240
PubMed Abstract | CrossRef Full Text | Google Scholar
Wiley, E. O. (1978). Det evolusjonære artsbegrepet revurderes. Syst. Biol. 27, 17–26. doi: 10.2307/2412809
CrossRef Full Text | Google Scholar
Wilson, E. O. (2017). Biodiversitetsforskning krever flere støvler på bakken. Nat. Ecol. Evol. 1, 1590–1591. doi: 10.1038 / s41559-017-0360-y
PubMed Abstract | CrossRef Full Text/Google Scholar
Yang, Zh, Og Rannala, B. (2014). Unguided arter avgrensning ved HJELP AV DNA-sekvensdata fra flere loci. Mol. Biol. Evol. 31, 3125–3135. doi: 10.1093/molbev/msu279
PubMed Abstract | CrossRef Full Text | Google Scholar
Yeates, D. K., Seago, A., Nelson, L., Cameron, S. L., Joseph, L., Og Trueman, W. H. (2011). Integrativ taksonomi eller iterativ taksonomi? Sys. Ent. 36, 209–217. doi: 10.1111/j.1365-3113.2010.00558.x
CrossRef Full Text | Google Scholar