XIX世紀の初めに、von Reckingausenは血管が細胞のシートで覆われていることを観察しました。 この1細胞の厚い層は、血液およびリンパ管の内部表面を覆う内皮細胞からなる。 ヒトでは、総内皮層表面は約35m2であり、約1-1,6 103個の内皮細胞が存在すると推定されている。 動脈、静脈、および毛細血管の全長は約90000kmです(世界中を2回旅行するよりも多く)。 内皮細胞は平坦であり、細胞接合部によって互いに接続されている。 それらの主な機能は、血液と他の組織との間の仲介者であり、正常組織および病理組織の両方における血液特性に寄与する。 それらはまた、血液と組織との間の免疫細胞の交換にとっても重要である。
- 1. 形態
- ソースと増殖
- 血管形成
- 血管形成
- 血管形成は、 成人では、内皮細胞の増殖率は非常に低い。 例えば、哺乳動物では、内皮細胞は、1ヶ月に1回分裂してもよく、または数年待つことができる。 これは、女性の生殖器官を除いて、正常な生理学下の組織では血管新生がまれであることを意味する。 しかし、それは組織の損傷や腫瘍のような病理学的プロセスで活性化することができます。 いくつかの物質は、内皮増殖因子(EGF)、酸性EGF、線維芽細胞増殖因子(FGF)、形質転換増殖因子(TGF)およびプロスタグランジンなどの血管新生を誘導する。 血管新生は、血管新生物質が周囲の組織から放出されるときに開始される。 近血管の内皮細胞は、通常、細静脈から、増殖し、新しい血管を形成し始め、他の内皮細胞をドラッグして、血管新生信号源に移動し始めます。 これらの内皮細胞は一時的にそれらの緊密な接続を失い、その結果、内皮細胞から放出される酵素によって迅速に消化される基底膜が明らかになる。 いくつかの内皮細胞は、増殖が続く間に消化された基底膜を通過する。 このすべてのプロセスは、血管新生信号が消失するまで、新しい血管の伸長および分岐を可能にする。 血管の先端は他の血管と融合して閉じた回路を形成し、死んだ終わりのない血液の流束を可能にする。
- 機能
- 血液の特性
- 免疫防御
1. 形態
内皮細胞は非常に平坦化された細胞であり、核も平坦化されていても、その核が細胞のより高い構造であるほどである。 細胞の形状は、内皮細胞が内張りしている管に適合している(図1、2、および3)。 より狭い毛細血管では、内皮細胞は血管の周囲のすべての細胞質を拡張してもよく、その結果、管は内皮細胞の列である。 より大きい管では、動脈および静脈のような、多くのendothelial細胞は管の全周囲を覆うために必要とされます。
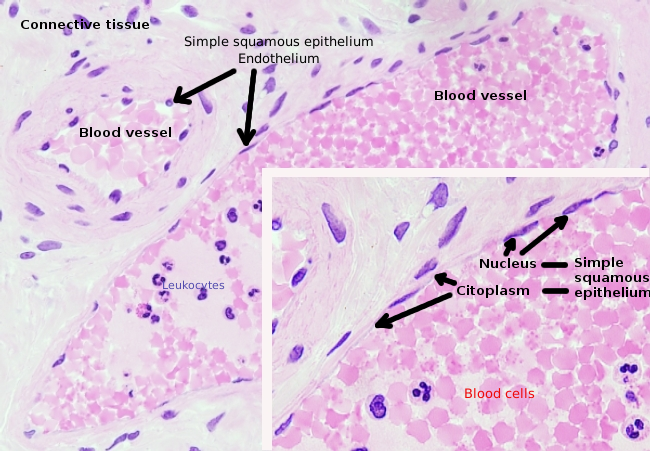
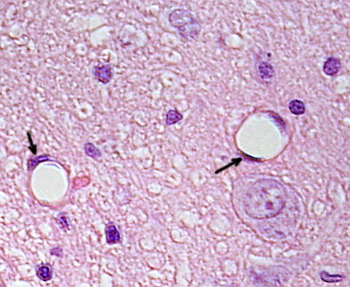
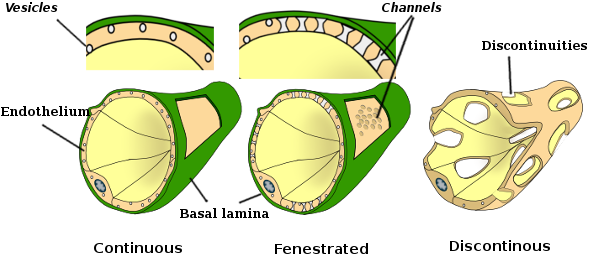
核周囲の細胞質は、ほとんどのミトコンドリアとゴルジ装置などの他の細胞小器官を含んでいますが、周辺の細胞質は薄く、小胞体は原形質膜の近くに見つけることができますが、いくつかの細胞小器官を持っています。 内皮細胞は多くの小胞を有し、その大部分は内細胞小胞である。 肝臓、腎臓皮質、内分泌腺などの一部の地域では、毛細血管の内皮細胞は、fenestratrionsとして知られている孔または小さな通路を示します。
それらは、直径が約60〜70nmの経路であり、約5nmの狭い通路であり、血液およびリンパを他の組織と直接通信する。 これらのタイプの内皮細胞を有する毛細血管は、有窓毛細血管として知られている。 細孔は群に分布し、その密度は内皮のタイプに依存する。 内皮細胞は、基底層と呼ばれる細胞外マトリックスの層によってsurronding組織から分離されています。
内皮細胞は、基底層と呼ばれる細胞外マトリックス 内皮細胞は、タンパク質ラミニン、フィブロネクチン、およびコラーゲンII型、IVおよびVでこの層に貢献します。
ソースと増殖
血管形成は、既存の血管 それは胚で起こります。 成人および胚においても、新しい血管、したがって新しい内皮細胞は、既に存在する血管から生成される。 このプロセスは、血管新生として知られています。 新しい内皮細胞は、血液およびリンパ管を分岐または分割することによって産生される。 血管新生は、成長している組織または哺乳動物の女性の子宮のような重いリモデリング下の組織、または腫瘍、炎症および創傷のような病理学において
血管形成
哺乳類の最初の内皮細胞は、胚の発生中に原腸形成後に生成されます。 それらは、血管芽細胞と呼ばれる細胞から分化し、群を形成し、短い血管に組織化される。 このプロセスは、最初に胚のビテリン嚢で起こる。 これらの初期の血管は成長し、互いに接続してネットワークを形成する。 その後、彼らは線維芽細胞と筋肉細胞を募集します。 この最初のネットワークは、次の開発段階で継続的な改造プロセスを経ます。 胚では、すべての内皮細胞は最初は類似しているが、後に静脈、動脈、毛細血管およびリンパ管内皮において分化する。 FGF2、BMP4、IHHおよびVEGFのようないくつかの分子は、分離された分化経路を誘導する。 Notch inducerは動脈内皮につながるが、その阻害は静脈内皮につながるため、非常に重要であると思われる。 リンパ内皮は枢機卿静脈内皮から形成され、COUPF-IIおよびSOX18の発現を必要とする。 さらに、内皮は、それが位置する器官に応じて分化される。 例えば、内皮は、血液脳関門を形成するために脳内にしっかりと密封された層を形成するが、血液との分子の交換を支持するために肝臓内で緩く組織化されている。内皮細胞が動脈および静脈管に組織化されるにつれて、いくつかの内皮細胞は、内皮-造血移行(EHT)として知られる分化プロセスによって造血細胞を生 このプロセスはまた妊娠の間に胎盤および他のティッシュの形成の間にのようなある大人のティッシュで起こります。
血管形成
血管形成は、既存の血管を分岐または分割することにより、新しい血管および内皮の形成である。
血管形成は、 成人では、内皮細胞の増殖率は非常に低い。 例えば、哺乳動物では、内皮細胞は、1ヶ月に1回分裂してもよく、または数年待つことができる。 これは、女性の生殖器官を除いて、正常な生理学下の組織では血管新生がまれであることを意味する。 しかし、それは組織の損傷や腫瘍のような病理学的プロセスで活性化することができます。 いくつかの物質は、内皮増殖因子(EGF)、酸性EGF、線維芽細胞増殖因子(FGF)、形質転換増殖因子(TGF)およびプロスタグランジンなどの血管新生を誘導する。 血管新生は、血管新生物質が周囲の組織から放出されるときに開始される。 近血管の内皮細胞は、通常、細静脈から、増殖し、新しい血管を形成し始め、他の内皮細胞をドラッグして、血管新生信号源に移動し始めます。 これらの内皮細胞は一時的にそれらの緊密な接続を失い、その結果、内皮細胞から放出される酵素によって迅速に消化される基底膜が明らかになる。 いくつかの内皮細胞は、増殖が続く間に消化された基底膜を通過する。 このすべてのプロセスは、血管新生信号が消失するまで、新しい血管の伸長および分岐を可能にする。 血管の先端は他の血管と融合して閉じた回路を形成し、死んだ終わりのない血液の流束を可能にする。
内皮細胞前駆細胞を有する領域は、背側大動脈および心内膜に見出されている。 間葉系細胞は、心内膜内皮細胞から分化することができ、三尖弁および心臓のいくつかの線維芽細胞を形成するが、心細胞は形成しない。 内皮細胞は外部シグナルに対して非常に賢明であり、内皮の完全性を維持することは、FGFのような他のシグナルによって媒介される能動的なプロセ 能動的なプロセスは、完全性のためだけでなく、細胞表現型を維持するためでもある。 内皮細胞が適切なシグナルを受信しない場合、それらはアポトーシスによって死ぬか、豊富な細胞外マトリックスを合成することができる間葉系細胞 内皮細胞から間葉系細胞への移行は、動脈硬化症および線維性心筋炎のようないくつかの疾患の背後にある。内皮はリンパ管と血管の受動的なライニングコートであるという考えを変更する必要があります。
機能
内皮はリンパ管と血管のパッシブライニングコートであるという考えを変更する必要があります。 内皮機能は多様であり、生物にとって不可欠である。 実際、それは心臓血管およびリンパ管の物理的構造であるが、それはまた、血管の内腔と周囲の組織との間の分子の交換を調節し、血液の生理学的特性 物理的な障壁のほかに、endothelial細胞は分泌の、新陳代謝および免疫機能を開発します。 それらは、成長因子、凝固剤および抗凝固剤、低密度リポタンパク質、一酸化窒素、セロトニン、エンケファリン、および他の多くのような分子の影響を受けて、その生理学的挙動を変化させる。 内皮細胞は、これらすべての物質に対する受容体を有する。 内皮細胞は、通常、血液と周囲の組織との間の障壁として作用する層を形成する。
内皮細胞は、血液と周囲の組織との間の障壁として 内皮細胞間の凝集は、タイトな接合部および付着性接合部などの細胞接合部によって達成される。 ギャップ接合も観察されているが、その主な機能は隣接する細胞間の通信である。 内皮細胞は、これらの癒着を調節し、バリアの透過性を変化させることができ、それは分子だけでなく、内皮を横断する細胞にも影響を及ぼし得る。 しかし、肝臓のようないくつかの器官では、内皮細胞ははるかに緩く包装され、内皮が障壁とはほとんどみなされないように多くの自由空間を残す。
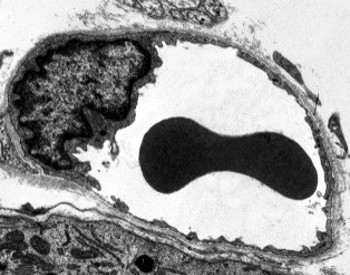
透過型電子顕微鏡画像は、内皮細胞の細胞質に多くの小胞を示す。 それらは、頂端(血液に面している)と基底外側膜(基底層に面している)との間の細胞内輸送に関与していると考えられている。 このタイプの輸送はtranscytosisとして知られています。 興味深いことに,経細胞は毛細血管の内皮細胞において,より大きな血管の内皮細胞よりも豊富であることが示された。 これは、毛細血管の内皮細胞は、血液と組織との間の分子のより強い交換を有するが、大径の血管では、それらは主に血液の伝導に関与することを示 有窓として知られているいくつかの内皮細胞は、周囲の組織と血液を直接接続する孔または非常に小さな管を有し、いくつかの小さなサイズの分子が内皮細胞の細胞質に入ることなく内皮を横断することを可能にする(図4)。 最後に、非常に透過性の内皮を有する器官がある。 肝臓では、シヌソイドは、内皮細胞が互いに自由空間を残す血管であり、バリアとしての機能はほとんど存在しない。
多くの細胞型は、体内の標的器官に向かって血液中を移動します。 そこでは、それらは血管の内皮を横断し、一般に毛細血管後静脈のレベルで交差する。 これは、内皮細胞が細胞が内皮層を通過させるために細胞接合部を改変しなければならないことを意味する。 白血球は、内皮細胞の頂端膜の特定の分子を認識し、固定することによって血管を出る。 セレクチン、インテグリンおよび免疫グロブリンは、内皮層への白血球の認識および接着に関与する。 セレクチンは、内皮表面上を転がる白血球の固定を開始する。 この初期接着は弱く、可逆的である。
多くの白血球は、炎症過程の間に血液を出て、冒された組織に移動する。 ケモキンは、損傷した組織によって放出され、内皮細胞のグリコカリックスにリンクされている白血球-魅力的な分子である。 ケモカインを有する内皮表面上を転がる白血球が活性化され,細胞-細胞接着が強くなり,白血球がその場に残る。 白血球の活性化は、内皮細胞の免疫グロブリンを認識するそのインテグリンの活性化をもたらす。 これらの免疫グロブリンは、ケモカインによる細胞の活性化後に内皮細胞膜に発現される。 細胞-細胞接着は内皮細胞中のカルシウム濃度を上昇させ、細胞接合部の解体および細胞質の収縮をもたらす。 このようにして、白血球は内皮細胞の境界に移動し、内皮を横断することができる。 接着分子はまた、白血球のこの動きに関与している。
血液の特性
内皮機能は、内皮を通過する分子や細胞を制御するだけではなく、より複雑です。 内皮細胞はまた、血圧、凝固、および他のいくつかの血液特性にも関与している。
原始循環系は600万年前に無脊椎動物に出現したと考えられているが、内皮を欠いていた。 内皮細胞は、より多くの層流束(乱流ではない)と、したがって、より効率的なガス交換を血液に提供する100万年前に登場しました。
内皮細胞は、血管の平滑筋に作用する物質を放出することによって血圧を調節する。 それらは、血管平滑筋を弛緩させる一酸化窒素(NO)およびプロスタサイクリンを放出する。 それらはまた血管の直径を減らすendothelinおよび血小板の活性化剤の要因を、両方解放します。 一酸化窒素はconstitutively解放され、適切な筋肉調子を提供しましたり、血小板の集合および白血球の付着を禁じます。 エンドセリンは強い血管収縮剤である。 内皮細胞がどの分子を放出しなければならないかをどのように決定するかはまだ明らかではないが、血液のフラックス特性を感じることができる頂端膜の機械受容体はもっともらしいかもしれない。
通常の条件下では、内皮細胞は適切な流動性を維持するのに役立つ分子を血液中に放出する。 流動性(抗凝固剤)と血小板凝集(抗血栓剤)の予防:彼らは2つのレベルで動作します。 蛋白質CおよびSは血の流動率に影響を与える重要な分子です。 Sタンパク質と複合体を形成するCタンパク質は、凝固因子VIIIaおよびVaを不活性化する。 Sタンパク質は内皮細胞によって合成される。 なお、endothelial glycocalixはトロンビンを不活性にすることができるヘパリンと同じようなglycosaminoglycanを含んでいます。
その抗凝固機能に関連して、内皮細胞は酸化窒素およびプロスタサイクリンを放出し、血小板中の環状AMPを上昇させ、血小板凝集をより困難にする。 これらの2つの分子は、血液中に連続的に放出される。 内皮細胞は、atpとADP、血小板凝集の両方の強力なプロモーターを除去し、その頂端膜にエクトヌクレアーゼを持っています。 さらに、内皮細胞は、プラミノーゲンをプラスミンに変換する活性化因子分子を放出し、血栓を除去することを好む。
これらの分子経路はすべて、内皮細胞が何らかのシグナルを受けたり、組織が損傷したりすると変化し、血液凝固および血小板凝集を招く可能性があります。
このような状況では、内皮細胞はその後、凝固および血栓症の能動的な参加者となる。
免疫防御
内皮細胞は、免疫防御において主要な役割を果たし、二つのメカニズムに関与している:tリンパ球への抗原提示と免疫細胞 マクロファージと一緒に、内皮細胞は、mhc-I(主要組織適合性複合体)を構成的に発現し、抗原提示に必要なMHC-IIを発現するように誘導される可能性があるため、tリンパ球に抗原を提示することができる。 内皮細胞は免疫記憶を活性化することができるが、新しいTリンパ球は活性化しない。 内皮細胞とTリンパ球との間には双方向の活性化があり,内皮細胞は炎症細胞を誘引するための分子を放出し,血液白血球を固定するための接着分子を発現する。
参考文献
Cines BD,Pollak ES,Buck CA,Loscalzo J,Zimmerman GA,McEver RP,Pober JS,Wick TM,Konkle BA,Schwartz BS,Barnathan ES,McCrae KR,Hug BA,Schmidt A-M,Stern DM1998. 生理学および病態生理学における内皮細胞血管障害の。 血液学のアメリカの社会のジャーナル。 91:3527-3561![]()
ファハルドLF. 1988. 内皮細胞の複雑さ。 臨床病理学のアメリカのジャーナル。 92:241-250.
Michiels C.2003. 内皮細胞が機能する。 細胞生理学のジャーナル。 196:430-443![]()