イントロダクション
国際保全政策は、生態系、種、遺伝の三つのレベルで生物多様性を認識し、管理は三つのすべてを保持することを目指すべきである(生物多様性条約、2007)。 これは、2010年に合意された生物多様性に関する条約”愛知生物多様性ターゲット”(https://www.cbd.int/sp/targets/)に明確に反映されており、生態系や種だけでなく、遺伝的多様性についても目標や目標に具体的な参照がある。 しかし、生物多様性の保全に対する現在のアプローチは、主に地理的地域、生態系、生態学的共同体、および種に基づいており、遺伝的多様性および種-個体群の連続性についてはあまり注目されていない。 実際、保全実務家の大多数は、集中的に管理されている絶滅危惧種内の種内単位を考慮して、生物多様性の指標として種に焦点を当てています。 この文脈では、単純な分類—名前付き種、および他の名前付き分類群(すなわち、亜種)、および時には種内の保全単位を採用する自然な欲求があります。 特に種は、多くの科学的および管理的努力を支える生物多様性の共通の尺度を提供する(Mace、2004;Wilson、2017)。 保全の基本的な単位として、種は、保護地域内の種のリスト、管轄区域内の絶滅危惧種の同定、およびバイオセキュリティ手順の基礎として、生物多様性 さらに、種は通常、私たちが保全問題を一般に伝える主体です。
自然の中で離散的な単位を単に識別するこの推進力は、いくつかの関連する現実に対抗します。 第一に、種分化は瞬間的ではない。 むしろ、それは典型的には長期化したプロセスである(Dynesius and Jansson,2014;Roux et al.,2016)は、おそらく異なる選択圧力の対象となる孤立した集団から、長い時間スケールで明確な種に進行し、環境変化または範囲としての単純な遺伝的合併によ ら、2 0 0 8;Rosenblum e t a l., 2012). 第二に、ほとんどの認識された種は遺伝的に均一ではありません。 発散(ドリフトおよび長期の単離)および網状化(geneflow、ハイブリダイゼーション、introgression、および組換え)は、種内の地理的およびゲノム変異を効果的に形成する(CutterおよびPayseur、2013;Edwards et al. いくつかの種は、長い孤立した集団に構造化することができ、そのうちのいくつかは完全ではあるがおそらく形態学的に不可解な種としての認識を保証することができる(Bickford et al.,2016)。 ることを示した。 第三に、認識されている限り、命名された種は、それらがどれほど進化的遺産を表すかという点で同等ではない—簡単に言えば、カモノハシ(古代に分岐した系統の代表者として)は、最近分岐したげっ歯類よりも価値があると考えることができる(Isaac et al., 2007). 第四に、種指定な鋳造石、その仮説をとして試験しなければならな新たなエビデンスのご利用になると(こんにちはet al., 2003). これらの問題のそれぞれは、一般的に種を区切ること、およびそれらまたはそれらの構成要素の集団に保全の注意を割り当てることに複雑さとあいまい さらに、種の境界はまた、異なる種の概念およびそれらを診断する方法の対象であり、これは保全生物学者の間で大きな驚きを引き起こした(Mace、2004;Frankham et al。 ることを示唆しています。
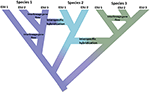
図1. 孤立した集団から明確な種に進行する連続体としての種分化は、異なる種を表す色との色のシフトによって示される。 個体群の分離のために生じるいくつかの系統は、種分化プロセスの開始を表す木の内部ノードを有する真の種に発展する。 他の系統は真の種に発達しないが、種が遺伝的に均一ではないことを示す時間の経過とともに発散する。 これらの系統は、進化的に有意な単位(Esu)として示されている。 系統間の遺伝子フローは、種1および種3について示されている。 この場合、同じ種に属する2つの系統が再現的に分離されていないため、進化的独立性は一時的である可能性があります。 それらは異なる系統(種)として残っているが、種間の遺伝子交換(色のシフト)が限られていると種間ハイブリダイゼーションが起こる可能性がある。
種と遺伝的多様性の両方を保護することを目的とした効果的な保全戦略の実施は、集団-種の連続体全体で種と他の単位をどのように区切る ゲノミクスの新しい機能は、これらの目標を達成するための機会と課題の両方を提供します。 以下では、次のことを考慮します: (i)ゲノムスケール解析を種の区切りと命名に対する保守的なアプローチとどのように調和させることができるか、(ii)ゲノムスクリーンによって提供される高解像度で、”種”を広範なinfra固有のカテゴリを含むように定義するいくつかの大陸にわたる既存の保全法をどのように使用できるか、(iii)ゲノミクスからの種分化プロセスと変異の構造化への洞察が法律とどのように一致し、その後管理にどのように一致するか。 次に、個体群から種を介して多様性の全範囲を保護するために、保全単位を立法的アプローチと調和させるためのいくつかの方法を提案する。
ゲノミクス、種分化、分類学
生物科学の多くの分野と同様に、系統学、分類群の命名の実践、およびそれらの関係とそれらがどのように形成されているかを理解することは、ゲノミクスと統計モデルによって大幅に進歩している。 全ゲノム配列決定を介するかどうか(例えば、Nater e t a l. ることができる(Snp、target capture−sequencing;Jones and Good、2 0 1 6)。; Leache and Oaks、2017)、私たちは今、遺伝的多様性のパターンの前例のない解決を持っています(Funk et al., 2012). ゲノムと種の相違の歴史を推論するためのますます強力な統計モデルと相まって、これらのデータは、種と遺伝的変異を生成する進化過程に新たな洞察を提供しています。
これらの新しいツールの適用は、二つの反対の洞察を生成します。 一方で、多くの研究は、命名された種、いわゆる不可解な種内の高度に発散した遺伝的集団を明らかにしている(Bickford et al. 2007年、Jorger and Schrödl、2013年; Striked et al., 2017). それは三十年のための生産的な努力となっている空間集団遺伝学と分子系統地理学の長い歴史の上に構築されているので、これは、新しいものでは, 1987). ゲノミクスがこの努力にもたらすのは、人口構造の解決の増加と、人口履歴を統計的に推測し、発散の代替モデルをテストするのに十分なデータの両方 一方、ゲノムスケールの解析では、(長い間植物で認識されていたように)動物種間の遺伝的交換(遺伝子導入)は、種分化の間と後の両方で、以前に考えられていたよりも一般的であり、新しい適応放射線を駆動することさえできることが明らかになっています(Mallet、2007;Rieseberg and Willis、2007;Arnold and Kunte、2017)図1。 まとめると、これらの洞察は、種分化が長期化することをさらに強調する(Roux e t a l. ら、2016)および系統地理学-系統発生学連続体を介して動作する反対の力を明らかにする(Edwards et al., 2016).
このすべてが分類学と保全の目的とどのように関連していますか? ゲノムスクリーンによってもたらされる解像度の増加は、種の横行した過剰分裂につながる可能性があるという強く正当な懸念がある(Isaac et al.、2004)、潜在的に管理の柔軟性を制限し、近親交配および最終的な絶滅に小さな遺伝的に発散する集団を委託する(Frankham et al., 2012). 逆に、Gippoliti e t a l. (2018)は、アフリカの有蹄動物の多くの種を認識する以前の失敗をもたらした分類学的慣性は、混合後の交配抑制と地理的に制限された分類群を保護するための失敗をもたらしたと主張している。 それらの見解(およびMorrison e t a l. 2009年)は、分類学的インフレが有蹄類の保存に悪影響を及ぼしているという証拠はないということである(Gippoliti et al., 2018). 関連する懸念は、不安定な分類法が保全の実践と公共の認識の両方を遅らせることである(Garnett and Christidis、2017)。 これらの懸念は、系統的生物学における種の概念と診断方法、特に種が最小限に診断可能な単位であることを保持する系統発生種の概念の適用(Cracraft、1983) しかし、分類学者の大部分は本質的に保守的であり、複数の証拠行にわたって一致がある場合にのみ種を命名する(Dayrat、2005;Padial et al. ら,2 0 1 0;Schlick−Steiner e t a l. 2010年:Yeates et al., 2011). 我々の見解では、種は、進化種の概念(Wiley、1978)と一般化された系統種の概念(de Queiroz、1998、2007)の意図に沿って、近親者と接触したときに大部分が無傷のままである堅牢な独立して進化する系統を表すべきであるということである。 実際には、これは、限られた遺伝的交換を可能にしながら、生殖分離または同じの合理的なサロゲートの直接的な証拠を必要とする(Singhal et al., 2018).
ゲノムデータは、種内および種間の分化および発散を同定するために以前よりも大きなパワーを提供し、これが分類単位の描写にどのように影響す 個体群が既に命名された種と比較して遺伝的にも表現型的にも分岐している場合、これらを別々の種として命名するための強力なケースがあります。 集団がゲノム分離がほとんどなく表現型的に強く発散している場合、それらの表現型の違いが(i)遺伝性であり、(ii)実質的な生殖分離を引き起こす可能性があるかどうかについての判断が必要である。 ゲノムスキャンは、制限された遺伝子の流れをテストすることによって、このような状況を解決することができます。 より困難な問題は、形態学的に類似しているが遺伝的に発散する集団が検出される場所である。 そのような集団は、多焦点データに統計的区切り法を適用することによって、別々の進化系統として容易に診断することができる(Carstens et al., 2013; しかし、それらは一時的な個体群か耐久性のある種ですか(Sukumaran and Knowles、2017)? 命名法の規則の範囲内で、DNA配列のみから種を認識し、配列ベースの診断を提供することは確かに可能である(Tautz e t a l. ら、2 0 0 3;Cook e t a l., 2010). 例えば、Murphy e t a l. (2015)は、明確な形態学的差異がないにもかかわらず、DNA配列データに基づいて六つのオーストラリアの砂漠の春の両脚類種を区切って記述する。 しかし、これは、システム主義者が「不可解な種」という用語をより一貫性と厳密さで適用する必要がある改善のための一般的な領域である(Struck et al., 2017).
DNA配列データが種の認識のための潜在的な重要な定義機能を提供する状況では、過度の分割を避けるために他の証拠を適用することが特に重要, 2015). 一つのアプローチは、そのような集団が自然接触ゾーンを形成する実質的な生殖隔離をテストすることです。 例えば、表現型的に区別できないカルリアトカゲの系統対の接触ゾーンの比較ゲノム解析では、遺伝的単離の指標は発散時間と強く相関していた(Singhal and Moritz,2013;Singhal and Bi,2017)(図2)。 これらの系統対のより発散的なものは、現在、正式には別々の種として認識されているが、接触ゾーンにわたる遺伝学的に広範な遺伝的交換の証拠を有するよ, 2018). これらの分類群における分岐と生殖分離との間の強い経験的関係を考えると、これらの著者は、上記で区切られたものよりもさらに高いゲノム分岐を有する(密接に関連する種の)二つのアロパトリック分離株と不可解分離株をさらに分離した。 この生殖分離を、深く発散しているが形態学的に類似した系統系統を有する他の同族種に拡張する(Potter et al. ら,2 0 1 6,2 0 1 8;Afonso Silva e t a l.,2017a)は、より多くの種が、特に島に記載されることが残っていることを示唆している(図2,Carlai amax and C. 実際、そのような種のいくつかは、相補的な表現型分析(Afonso Silva e t a l.,Nucleic Acids Res.,Nucleic Acids Res.、2017b)。 より最近の相違を持つ他の系統は、核遺伝子の大規模なセットを使用して別々の系統として統計的に区切られているが、種内の進化的に有意な単位(Esu)
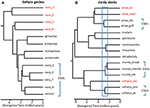
オーストラリアのモンスーン熱帯地方(AMT)からトカゲの種の境界の系統発生論的解決。 いずれの場合も、関係と相対的な発散時間は、Starbeast2で実装された複数種の合体を使用して100エクソンから推定されますが、二つの例の間の時間スケールは必ずしも同じではありません。 (A)Gehyra geckosのnana群の結果(Moritz e t a l., 2018). G.nana内の系統の系統発生の多様性は、以前に認識されているように、すでに記載されている他の四つの形態学的に特徴的な種を含んでいた。 範囲の重複および形態のその後の分析は、最も系統発生的に発散する系統(赤色の分類群標識)のそれぞれを別々の種として認識することを導いた(Doughty et al., 2018). 現在認識されているように、G.nanaに残っている(形態学的に不可分な)系統は、Esuとみなすことができる。 (B)AMTからCarlia skinksにおける種の境界の継続的な解決。 いくつかの系統は、遺伝種の区切りと診断形態学的差異(C.insularisおよびC.isostriacantha)の事後同定の組み合わせに基づいて、すでに種として認識されている; アフォンソ-シルバ他、2017b)。 他の錯体については(Potter e t a l. 2016年、2018年)、分類学的改訂が進行中である。 青色の長方形は、発散と相関する実質的な生殖分離が、関連種の形態学的に不可解な系統間の接触ゾーンで観察された発散時間の範囲を示す(Singhal And Bi、2017)。 保存的には、および以下(Singhal e t a l.、2018)、我々は候補種として、この経験的閾値よりも大きな発散を有する三つの遺伝系統を認識する。 これにより、Carlia amax、Carlia munda、Carlia rufilatus(青い括弧)のそれぞれに遺伝的に異なるEsuが残される。 スケールバーは、数百万年での発散時間の推定値を表します。極端な場合には、ゲノム法は、単一の分類学的種が遺伝的に異質であるだけでなく、系統発生的な意味で他の名前の種も含むことを明らかにする。
このような”パラフィリー”は、遺伝的に細分化された単一の種からの最近の種分化によって生じる可能性がある(Patton and Smith、1994)が、遺伝的に異質な分類群が複数の種を表す可能性があるという仮説も生成する。 再び、より多くの証拠が必要です。 例として、オーストラリア北部のゲヒラヤモリトカゲの複合体の系統解析は、9つの進化系統を区切っており、それらの間には既に命名された4つの種が系統発生的に埋め込まれている(Moritz et al.,2018;図2). その後の地理的重複(系統学的分離を保持しながら)と微妙な形態学的差異の証拠は、四つの最も系統的に発散する系統の命名を支持した(Doughty et al., 2018). 逆に、より密接に関連した(まだ統計的に分離可能な)系統は、重複する分布ではなく隣接していたが、形態学的に診断可能ではなかったため、複数のEsuを これらのEsuの境界を越えたサンプルのその後のゲノムスキャンは、少なくともいくつかは、種を認識するために取られた元の、保守的なアプローチを検証し、実質的な遺伝的交換を持っていないことを明らかにしました。
他のケースでは、植物ゲノム研究は、Snpの単純な序列によっても、集団サンプル間でスタークで予期しない相違を明らかにすることができ、診断表現型の特性を分析することができる不可解な分類群の存在を示唆している。 例えば、集団間のゲノムの発散とユーカリサルブリスのエコタイプの違いのためのSNPベースの証拠は、エコタイプの適応は、系統間の遺伝子の流れを最小化する上で重要な要因であると考えられて、地理的な重複の領域で彼らの独自性を維持し、二つの異なる分子系統を明らかにした。 (Steane e t a l.,2015;図3). 別のケースでは、オーストラリアのペラルゴニウムでは、ゲノム分析は、形態学的および生殖的研究と組み合わせて、特に以前に使用された形態学的形質が不安定で系統発生と不和であることが判明している分類学の有意な変化をもたらした(Nicotra et al., 2016).
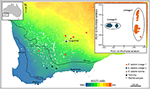
図3. SNPデータを用いたゲノム解析は、広範なユーカリサルブリスにおける不可解な系統を同定した。 各系統のサンプル個体群の位置は、西オーストラリア州南西部の乾燥指数勾配にわたって示されている。 16,122DArTseqマーカーから派生したバイナリ遺伝的距離に基づくPCoAは、二つの異なる系統を示しています。 Steane et al. (2015).
上記のケースは、統合的分類法で使用されている漸進的仮説検定アプローチを示しています(Padial et al. ら,2 0 1 0;Yeates e t a l., 2011). さらに、これらのケースは、独立した遺伝子の1000年代から高品質のデータを得るためにゲノム削減シーケンスのいくつかのフォームを使用して、結果にはるかに大きな自信を与えて、ちょうど色素体遺伝子座またはいくつかの核遺伝子を使用している場合よりも。 統合的な分類法を適用することで、多様性の認識を劇的に変えることができる方法の例は、既知の種の多様性をほぼ倍増させたマダガスカルのカエルの遺伝的、音響的および形態学的調査から来ている(Vieites et al., 2009). これらの著者らは、分類学的解像度と合同性の状態を反映した用語のシリーズを提案した: 遺伝的および表現型的に発散するが、まだ分類群に名前が付けられていないための確認された候補種、形態および音響がまだ検討されていなかった遺伝的に発散する分類群の未確認の候補種、および遺伝的に発散しているが表現型的に不可解であることが知られているエンティティのための深く同種の系統。 ゲノム解析は、これらの分類群にはまだ適用されていないが、確かに候補種対同種系統についての決定を知らせるだろう。
ゲノムアプローチは、保存的に適用された場合でも、間違いなく形態学的に保存的分類群のさらなる分裂につながるでしょう。
しかし、逆に、同じアプローチが過剰に分割された種または誤って診断された種を明らかにする明確な例がある(Nicotra et al., 2016). もちろん、小規模なデータセットに基づいて、植物や動物種の複合体に関する多くの分子研究がすでにあります(例えば、 種が誤診された場所、または複数の種が分類学的に外接されているが、現在は単一の種であると考えられている場所を同定した。 例えば、Rabosky e t a l. (2014)は、ミトコンドリアと核遺伝子の系統発生を使用してscincid Ctenotus inornatus複合体を修正し、種の純減少と種の境界の再配置をもたらした。 植物では、最もよく知られた例のいくつかは、ヨーロッパの蘭属Ophrysとオーストラリアの蘭属Caladeniaなどの蘭で発見されており、分子研究は、形態学的形質および偽繁殖受粉症候群に基づくものよりも、種として認識される実体が有意に少ないことを示している(Devey et al. ら、2 0 0 8;Swarts e t a l., 2014).
ゲノムスケールデータは、表現型の並列解析で保存的かつ理想的に解釈されると、特に形態学的に保存的な分類群において、分類学的変化をもたらし続, 2018). しかし、一部の人が表明した懸念とは対照的に(Garnett and Christidis、2017)、たとえ結果が一時的に不便であっても、これは進化系の適切な追求であると考えています。
ゲノミクスと種内保全単位
分類学的区切りや種レベルの議論よりも論争は少ないが、多くの点で複雑であるが、保全行動に考慮する必要がある種内多様性の重要な要素としての保全単位の定義と区切りについて重要な議論が行われている(Ryder、1986;Waples、1991;Dizon et al. ら、1 9 9 2;Moritz、1 9 9 4;Voglerおよびd e Salle、1 9 9 4;Crandallら、1 9 9 4;Mol. ら,2 0 0 0;Fraser and Bernatchez,2 0 0 1;Avise,2 0 0 4;Funk e t a l.,2012)(図4). 我々が上記で提唱しているゲノム証拠から種を認識するための慎重なアプローチを考えると(そしてOliver et al. ら、2 0 1 5;Singhal e t a l. いくつかの分類学的種は、依然としてそれらの中に長い孤立した集団を含むことは避けられない。 進化的に有意な単位(Esu)または指定可能な単位(Du)のような概念は、既存の分類学を補完するために提案されている(Ryder,1 9 8 6;Waples,1 9 9 1;Moritz,1 9 9 4;Mee e t a l., 2015). 種と同様に、解釈および基準は様々であるが、共通の意図は、保全行動のための種内多様性の主要な要素を認識することである(Funk et al., 2012). Moritz(1994,2002)は、種内の独立して進化するセグメントを識別するための単純ではあるが実用的な分子アプローチ、多様性の”歴史的”軸を提唱した。 ら、2 0 0 0;Fraser and Bernatchez,2 0 0 1;Waples e t a l. ら,2 0 0 1;Meeら,2 0 0 2;Me e t a l.,2015)(図4). これらの選択肢の相対的な利点が何であれ、集団ゲノミクスの技術的および分析的ツールが、種内多様性の両方の成分を堅牢に同定できることは明 ら、2 0 1 2;Hoban e t a l. ることができます。
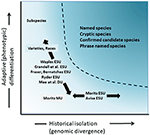
図4. 遺伝的多様性の2つの要素への分離: 自然選択を介して発生する適応変異、および表現型と歴史的な分離またはゲノムの発散を介して評価中立発散の分析かかわらずアッセイ。 曲線の上の暗い影付きの領域は、ほとんどの概念の下で個体群が別々の種として考慮される可能性が高い条件を示しています。 様々な保存単位の概念と以下に固有の分類学的カテゴリは、遺伝的多様性の二つの軸に関連して示されている(Moritz、2002の後)。これは質問につながります:なぜEsuを亜種として名前を付けるだけではないのですか? 亜種は伝統的に定義されており、植物学者と動物学者の両方によって表現型が異なる、地理的境界で相互に相互作用する可能性のあるアロパトリックセッ そのため、必ずしも歴史的に孤立していない適応的な発散を反映する可能性があります。 亜種の指定は、植物において広く使用されており、地理的および生態学的な違いは、それらが区別される主要な手段である(Hamilton and Reichard、1992)。 これまでのところ、亜種は、進行中の遺伝子の流れの有無にかかわらず、ローカル適応を明示するように、我々は表現型で定義された亜種は、保全に認識され、Esuと
集団ゲノミクスと同じように、中立遺伝子座と適応遺伝子座の両方を利用することにより、Esuの同定においてはるかに高い精度を提供し、制限された遺伝子フローを特徴とする人口統計的に独立した単位として管理単位(Mu)をより容易に区切るために多数の中立遺伝子座を利用する手段を提供する(Funk et al., 2012). ここでの焦点の大部分は、保全のための主要な特定内単位として認識され、いくつかの管轄区域の下で保護のために受け入れられているESUおよびdusのような類似の単位にあるが、MUsは人口学的に独立した集団を表すことも同定されている(Palsbøll et al. より大きなESUを保持するために管理することができる(Moritz,1 9 9 4;Funk e t a l.,2 0 0 7)。, 2012). 管理単位は、種の個体群を監視および管理する上で重要であり、特に漁業管理に関連する「在庫」とも呼ばれている(Avise、2004;Laikre et al., 2005). Moritz(1994)によるMUsの定義は、「対立遺伝子の系統発生的識別性にかかわらず、核またはミトコンドリア遺伝子座における対立遺伝子頻度の有意な発散を有する集団」として一般的に広く受け入れられているが、Palsbøllらによって指摘されているように。 (2007)では、人口統計学的結合性は、歴史的な遺伝子フローのレベルではなく重要な要因である。
種内多様性の重要な要素として保全単位を区切ることは、遺伝子流の回復または増強による小さな近交系集団の遺伝的救助にも重要な意味を持ち、集団絶滅の増加を回避するためには、遺伝的救助をより広く考慮する必要があることがますます明らかになっている(Love Stowell et al. ら、2 0 1 7;Ralls e t a l., 2018). Moritz(1999)によって、EsuではなくMUsを混合することは遺伝的救助のための適切な戦略と考えることができることが提案された。 Frankham et al.によって強調されているように。 (2012)分類学的に認識された種の混合は、一般的に管理者によって許容されるとはみなされず、その場合、分類学的な過剰分割は、小さな近交系集団を救 我々は、MUsなどの種内ユニットを区切るために、より厳密なアプローチは、この問題を軽減することができるが、それは移動速度と適応発散のゲノム推定は、種の境界と種内変異を定義するだけでなく、遺伝的救助戦略を設計するだけでなく、はるかに堅牢なガイダンスを提供する可能性を秘めていることも明らかであることを示唆している。
系統発生学と保全
系統発生学的方法と組み合わせたゲノミクスは、容易に分類群間の関係と進化の距離を解決するだけでなく、種を区 これは、遺伝子ツリーの不均一性を説明し、先祖の多型による先端長の過剰膨張を避けるため、連結ではなく、種ツリー法を使用して最もよく達成される(Edwards et al. ら、2 0 1 6;Ogilvieら、2 0 1 7;Smith e t a l., 2016). 多数の遺伝子座が必須ではないことが多いが、より多くの遺伝子座を含むことは、しばしば、関係および分岐長の両方の解決を改善する(例えば、Blom et al., 2016). 保全の重要性は二重です。 第一に、系統発生評価は、それらが記述されているかどうかだけでなく、種またはEsuの独自性を考慮することを可能にする。 従って、EDGEプログラム(Isaac e t a l.,2007)は、通常のIUCN基準に加えて、それらがどれだけユニークな進化の歴史を表しているかに基づいて、保全のための種の重み付けを提唱しています。 第二に、枝の長さ自体は、分類学とは無関係に生物多様性の価値を表すことができ(Faith、1992)、これは保全のための地域の優先順位付けに価値を示している(Laity et al. ら,2 0 1 5;Rosauerら,2 0 1 5;Rosauerら,, 2016, 2018). この意味で、系統発生多様性や系統発生固有性などの尺度を用いた系統発生指標を採用することにより、保護の対象となる進化的多様性の高い領域を強調し、生物多様性保全の改善のための計画プロセスにおいて高い優先順位を与えることが可能になる。 このアプローチは、地理的スケールの範囲にわたって生物の多様なグループのための遺伝的多様性と進化過程の保護を強化することができます(Laity et al.,2015)ゲノムエビデンスが種または特定単位内の認識にどのように翻訳されるかの変化に対して、より堅牢な優先順位を付ける領域についての決定を
現実世界—立法の定義
生物多様性保全のための政策と立法の枠組みは、国際条約と戦略から、国家戦略と法律、国の州または地域の法律と 生物保全に関する主要な国際的な文書には、締約国会議(COP)、世界遺産条約、世界生物多様性戦略、モントリオール森林保全プロセス、湿地に関するラムサール条約、渡り鳥種に関する条約、絶滅危惧種の国際取引に関する条約(CITES)、国際自然保護連合(IUCN)絶滅危惧種レッドリスト、国連気候変動枠組条約などがある。 これらの憲章はすべて、生物多様性保全の基本的な目標として種と遺伝的多様性を保全することの原則を具現化しており、種は保護のために広く定義されている単位である。 しかし、法律や国際指令の下での種の定義は非常に可変です。 それらは、種および種内分類群の分類学的認識を組み込むが、しばしば「種」の法的定義の下でカバーされる遺伝的/進化的概念も組み込むことができる(表1)。テーブル1

リストと保護のための”種”を定義する世界の法律と指令。
保護できる広範なインフラスペシフィックレベル(表1)を考えると、国の法律と関連する上場プロセスは、遺伝的多様性の保全を促進すべきであ 例えば、世界中の法律を考えるときの「種」の定義(表1)には、次のようなものがあります。: 種、亜種、品種、および地理的および/または遺伝的に異なる集団、南アフリカの法律の下では、品種、地理的人種、系統および雑種も含むことができる。 したがって、世界的なほとんどの法律の下では、”種”は、より広範な種の分布の一部であり、個体群-種の連続体を様々な程度に反映する様々なinfra-specificレベルを反映 これは、分類学者、進化生物学者、保存遺伝学者が、この連続体内の単位を指定するためのより一貫した均一な方法を見つける際に直面する避けられない難しさと平行しています。
いくつかは、種とインフラ固有の不確実性の政策的影響に関する懸念が見当違いであると主張している(Hey et al.ここでは、このような議論は、生物学ここんにちは、生物学的には、,2003),しかし、我々はまだ法律がレベルの範囲で遺伝的多様性を保護するためのメカニズムを提供することができますが、責任は、これらのレベルが種から個々の集団に至るまでどのように区切られるべきかについてより明確かつ一貫性を提供するために生物学者にあるという見解のままである。 現在、管理者や議員は、集団種の連続体全体の単位を区切るときに、科学者によって提供される種の概念とインフラ固有の単位の途方もない配列を残 例えば、すべての管轄区域では、分類学者によって認識された種を保全の重要な単位として考慮しています(表1)が、種の概念に関する分類学者の間ではまだ広範な議論があり、種が実際にどのように診断されるかはグループ内またはグループ間で異なる。 これは、分類学に対するより規制されたアプローチ(Garnett and Christidis、2017)の最近の呼び出しにつながっており、他の人は分類学の規制が答えではないと反論している(Thomson et al., 2018). 私たちはThomsonらによって作られた点のいくつかに同意しますが。 (2018)我々は、名前によって暗示される特定の分類群概念を明示的に参照するために、法律自体を変更する必要があることを提案するのはむしろ素朴であることを提案することを提案することを提案することを提案することを提案する。 これらおよび以前のこのような議論は、進化生物学者および分類学者が、ゲノムおよびその他の証拠を種または種内単位の境界にどのように翻訳するか、およびこれらの分類群が保全および保護のための関連する法律にどのように関連するかについての明快さ(透明性)を確保することの必要性を強調している。
管理への翻訳
上記のように、すべての法律および指令は、種レベルを保全の基本単位として認識するだけでなく、様々なインフラ固有の単位の認 多くはまた、保全の優先順位付けと正式な保護のために考慮されたときに種の正式な命名と分類学的記述のための暗黙の要件を持っています。 例えば、IUCNレッドリストガイドラインの下では、名前のない種のリストは推奨されておらず、新しい種の記述がiucnレッドリストに含まれてから4年以内に公表されるか、または削除されるという要件がある。 これは、種および特定のレベルの両方での実体の分類学的命名および記述の重要性、多くの植物および動物群の限られた分類学的情報(Mace、2004;Oliver and Lee、2010;Wilson、2017)、およ ら、2 0 1 3;Wege e t a l. ら、2015年、Wilson、2017年)。
分類学的に命名された種および亜種および品種などの種内分類学的カテゴリーは、一般的に正式なリスト、保全の優先順位付けおよび保護のための基本的な単位を提供するが、いくつかの管轄区域では種内保全単位の他のカテゴリーが正式に認識されている。 これらは一般に、ゲノムデータと表現型データを組み合わせて同定される。 たとえば、米国絶滅危惧種法は、esuに広く基づいている個別の個体群セグメント(DPSs)を認識しています(Waples、1991)、カナダの種リスク法はDUsを認識しています。 カナダ種リスク法の下での「異なる集団」の運用上の解釈では、DUsは、種の進化的遺産の重要な構成要素を表し、自然分散によって置き換えられそうにな, 2015).duとDPSsの両方が概念的にEsuと重複しています(Waples et al. ら、2013)、北西米国サケ科(Waples、1995)およびカナダのコレゴン科(Meeら、2013)を含む北米の魚に最も広範囲に適用されている(Meeら、2013)。 興味深いことに、米国のDPSsは脊椎動物にのみ適用され、カナダではDusはその行為の下に記載されている任意の生物に適用することができます(Waples et al.、2013)、dusは現在節足動物、軟体動物、および植物のために記載されています。 オーストラリアでは、カナダと同様に、1999年の環境保護および生物多様性保全法(EPBC法)の下で認識されている生物については、DUSsやDPSsなどの具体的な名前のカ しかし、今日まで、彼らは脊椎動物のためにのみ認識されています。 米国では保全のためにリストされている127脊椎動物Dpsがあります(https://ecos.fws.gov/ecp/);カナダでは70DUsはすべての生物をカバーしてリストされています(http://www.registrelep-sararegistry.gc.ca/species/schedules_e.cfm?id=1); また、オーストラリアでは、哺乳類、魚類、鳥類を含む7種の脊椎動物の個体群がEPBC法(http://www.environment.gov.au/cgi-bin/sprat/public/publicthreatenedlist.pl)の下で特定の保全状況を有すると認識されている。 米国およびカナダにおけるこれらの集団の評価は、一般的に遺伝情報を使用するが(Waples et al. 2013)、オーストラリアでは、それらは通常、地理的および管轄の同定に基づいているが、これらのケースのいくつかでは、それらは哺乳動物の遺伝的分化の分析に ら、1 9 9 9;Armstrong、2 0 0 6)およびfish(ColganおよびPaxton、1 9 9 7;Ahonenら、1 9 9 8)。, 2009).
カナダ(DUs)の植物および無脊椎動物のためのinfra-specific conservation unitsのリストではなく、米国またはオーストラリアでは、conservation unitsの境界に特に関連していないが、生物多様性 特定の生物の集団のこの優先的なリストは、保全単位を区切る科学とはあまり関係がなく、一般の意識、政治的便宜、リストされる生物のカリスマ性に基づ, 2013).
オーストラリアでは、よく知られている哺乳類、鳥類、魚類の一部の個体群がEPBC法の下で保護されていますが、爬虫類、無脊椎動物、植物の分子研究に基づ 例えば、ESUの要件に容易に適合する種内の高度に構造化された集団および深く発散する系統の顕著な例は、熱帯地方の低分散脊椎動物の範囲(図2) 同様に、両生類やヤスデのようなオーストラリアの無脊椎動物では、重要な遺伝的構造、および系統地理学的パターンは、特定の生息地に限定され、少数の集団 ら,2 0 0 7;Murphy e t a l. ら、2 0 1 3;Nistelberger e t a l.、2014a)。 しばしば、これらの短距離風土病は、保存上の懸念の分類群である(Harvey e t a l. ら、2 0 1 1)、および固有病ホットスポットに共存している(例えば、Oliver e t a l. ら、2017)、それ自体が多分類群保存の注意のための高い優先順位である(Rosauer et al.、2018)、キンバリー地域、および南西西オーストラリア州のスターリング山脈など。
オーストラリアの植物では、分離した歴史的に孤立した個体群システム、地質学的および地質学的複雑さを反映し、植生および気候の面で対照的な生息地を占有する異なる属の種の範囲内の異なる個体群群を特定する集団遺伝学的および系統地理学的研究もかなりの数ある(Broadhurst et al., 2017). 遺伝的に異なる集団は、西オーストラリア州南西部からの種において特に明らかである(Coates,2000;Byrne et al. ら,2 0 1 4)、アカシア(Mimosaceae;Coates,1 9 8 8)、Atriplex(Amaranthaceae;Shepherd e t a l.,2 0 1 4)、およびAmaranthaceae(Amaranthaceae;Shepherd e t a l. ら、2 0 1 5)、Banksia(Proteaceae;Coates e t a l. ら、2 0 1 5)、Lambertia(Proteaceae;Byrne e t a l. ら、2 0 0 2)、Hakea(Proteaceae;Sampson e t a l. ら、2 0 1 4)、Eremophila(Scrophulariaceae;Llorens e t a l.,2015),Pultanaea(Fabaceae; Millar and Byrne,2 0 1 3)、およびCalothamnus(Myrtaceae;Nistelberger e t a l.、2014b)。 多くの場合、これらの個体群は州政府機関による保全計画において認識され、環境影響評価において考慮される(Shepherd et al. 2015年には、法律の下で保護のために記載されていないにもかかわらず、。 しかし、我々は、これらのユニットの正式な認識は、長期的にはその保全を改善することを認識しています。
これらのオーストラリアの例は、種レベル以下の遺伝的多様性のこの層を認識する必要性が高まっていることを強調しています。 課題は、保全の関心のための競合する要求に直面して、保全の懸念である高度に構造化された集団と深く発散系統が保護されていることを確認する 象徴的な種を超えて、この挑戦はカナダを除いて世界的にも明らかです。 明確に描写された保全単位を保護しないと、不可解な絶滅のリスクと重要な遺伝的多様性の喪失が増加する可能性が高く、多くの場合、何百万年もの間種の中に存続してきた系統や歴史的に孤立した集団が関与する可能性がある(Moritz、2002)。
結論と勧告
絶滅危惧種の法律またはIUCNなどの国際機関の下で種と様々なinfra固有のカテゴリのリストは、種の集団連続体全体の遺伝的多様性 しかし、生物多様性保全への現在のアプローチは、主に遺伝的多様性と以下の特異的変異に与えられたあまり注意を払って、この連続体の種の終わりに焦点を当てています。 種内レベルの遺伝的変異に対するこのより限定された焦点は、保全管理者が管理行動および優先順位付けにおいて遺伝情報を明示的に考慮する ら、2 0 0 8;Laikre e t a l. ら,2 0 1 0;CookおよびSgrut,2 0 1 7;Taylor e t a l., 2017). 科学と保全の実践の間のこのギャップは、遺伝的多様性を保存し、管理するためのより効果的なグローバルなアプローチを妨げます。
私たちは、保全の単位に対する生物多様性の科学者からの明確さの欠如は、主に二つのレベルでの不一致に基づいていることを示唆しています。
最初は、複数の種の概念と分類学的グループ間で矛盾を作成する種を区切るために使用される様々な診断方法に関係しています。 第二は、分類学者、進化生物学者、保全生物学者による、特定の分類群やEsu、DUs、MUsなどの他の実体をどのように扱うかについての一貫性と合意の欠如に関す
立法は、分類群(または”種”)がどのように定義されているかという点ですでに十分に柔軟であることが多いと考えているが、集団種の連続体に体系的に対処する必要があると主張し、ESUのような概念に与えられたより大きな認識を持つ保全のための単位をより明確に定義し、これらが関連する法律とどのように関連するかを議論する。 我々は、この問題は間違いなく、徐々に微細なスケールで分化を検出するためのゲノミクスの力と種とinfra特異的エンティティを区切るためにその増加使
前の分析から、我々は以下の推奨事項を提供しています:
(i)種を区切るときにゲノムデータは、通常、遺伝データだけを超えた証拠を含む統合的な分類学的アプローチを使用して、保存的に解釈されるべきである。
(ii)遺伝学者が種内の複数の系統または異なる集団を特定し、これらが種レベルへの昇格を保証せず、分類学的に認識されていない場合、保全状態または影響評価の実用的な評価のために明確に正当化された種内単位を特定するためのより強力な推進力とより均一なアプローチがあり、これらがその管轄に関連する法律の文脈において保全のための単位として明示的に認識されること。
(iii)Iucnは、保全遺伝学専門家グループおよび地域サブグループを通じて、最良の科学的実践に合わせた種内ユニットを認識およびラベ
(iv)種の進化的ゲノミクスと個体群から種への連続から出てくるより微妙な見解は、科学者によって保全管理者や政策立案者によ
より広義には、最近多くのレビューで行われた勧告をエコーします(Frankham、2010;Santamaría and Méndez、2012;Hoban et al。 ら、2 0 1 3;Moritz and Potter、2 0 1 3;Laity e t a l. ら、2 0 1 5;Pierson e t a l. ら,2 0 1 6;CookおよびSgrut,2 0 1 7;Taylor e t a l. 分子系統学、分子分類学、系統地理学、集団遺伝学研究からの成果は、遺伝的多様性の保存と進化過程の管理に関する重要な情報を提供できることを、科学者が政策立案者、政府機関、非政府組織、および実務家とのコミュニケーションを改善し、意識を高める必要性を強調する(2017)。
著者の貢献
DC、MB、CMはすべて原稿の準備と改訂に貢献しました。
利益相反に関する声明
著者らは、この研究は、利益相反の可能性と解釈される可能性のある商業的または財政的関係がない場合に行われたと宣言している。
謝辞
このプロジェクトは、オーストラリア研究評議会(ARC Linkage Grant No. LP120200063)、オーストラリア国立大学と生物多様性、保全、および観光の西オーストラリア省。
Afonso Silva,A.C.,Bragg,J.G.,Potter,S.,Fernandes,C.,Coelho,M.M.,And Moritz,C.(2017a).この記事は、ウィキペディアのコエーリョ(改訂履歴)の記事を複製、再配布したものにあたり、 熱帯専門家対気候ジェネラリスト:オーストラリア北西部からのCarlia skinksの姉妹種の多様化と人口統計学的歴史。 モル エコール 26:4045–4058. 土井:10.1111/mec.14185
CrossRef全文|Google Scholar
Afonso Silva,A.C.,Santos,N.,Ogilvie,H.A.,And Moritz,C.(2017b). 複数種の合体法と形態を持つ二つの新しい北西オーストラリアのレインボースキンクスの検証と記述。 PeerJ5:e3724. ドイ:10.7717/peerj.3724
PubMed Abstract|CrossRef全文|Google Scholar
Ahonen,H. ら、harcourt,R.G.,およびStow,A.J.(2 0 0 9)。 核とミトコンドリアDNAは、imperilled灰色の看護師のサメの個体群(Carcharias taurus)の分離を明らかにする。 モル エコール 18, 4409–4421. doi:10.1111/j.1365-294X.2009.04377.x
Pubmed Abstract|CrossRef Full Text|Google Scholar
Armstrong,K.N.(2006). Rhinonicteris aurantia(Chiroptera:Hipposideridae)の系統地理学的構造:保全への影響。 Acta Chiropterologica8,63-81. 土井: 10.3161/1733-5329(2006)82.0.CO
CrossRef全文|Google Scholar
アーノルド、M.L. およびKunte,K.(2017)。 適応遺伝子交換:混和と進化の革新のもつれた歴史。 トレンド-エコール… エヴォル 32, 601–611. 土井:10.1016/j。2017.05.007
PubMed Abstract|CrossRef Full Text|Google Scholar
Avise,J.C.(2004). 分子マーカー、自然史、および進化。 2Nd Edn. オックスフォード大学出版局(オックスフォード大学出版局)。
Google Scholar
Avise,J.C.,Arnold,J.,Ball,R.M.,Bermingham,E.,Lamb,T.,Neigel,J.E.,et al. (1987). 種内系統地理学: ミトコンドリアDNAは、集団遺伝学と系統学の間の橋渡しをしています。 アン エコールレヴュー シスオ… 18, 489–522. doi:10.1146/annurev.es.18.110187.002421
CrossRef全文/Google Scholar
Bickford,D.,Lohman,D.J.,Sodhi,N.S,Ng,P.K.,Meier,R.,Winker,K.,et al. (2007). 多様性と保全の窓としての不可解な種。 トレンド-エコール… エヴォル 22, 148–155. 土井:10.1016/j。2006.11.004
Pubmed Abstract|CrossRef Full Text|Google Scholar
Blom,M.P.K.,Bragg,J.G.,Potter,S.,And Moritz,C.(2016). 遺伝子ツリー推定の不確実性のための会計処理:要約-オーストラリアのトカゲの挑戦的な放射線における合体種ツリー推論。 シスト バイオル 66, 352–366. doi:10.1093/sysbio/syw089
PubMed Abstract|CrossRef Full Text/Google Scholar
Broadhurst,L.,Breed,M.,Lowe,A.,Bragg,J.,Catullo,R.,Coates,D.J.,et al. (2017). オーストラリアの植物相の遺伝的多様性と構造。 ダイバーズ ディストリブ 23, 41–52. doi:10.1111/ddi。12505
CrossRef全文|Google Scholar
Byrne,M.,Coates,D.J.,Forest,F.,Hopper,S.D. ら、krauss,S. (2014). “A diverse flora-species and genetic relations,”In Plant Life on the Sandplains in Southwest Australia,A Global Biodiversity Hotspot,editors H.Lambers(Crawley,WA:University of Western Australia Publishing),81-99.PubMed Abstract
Byrne,M.,Macdonald,B.,And Coates,D.J.(2002). アカシアacuminata(マメ科:Mimosoideae)内の系統地理学的パターンと高いcpDNAの多様性。 西オーストラリア州の複合体。 J.Evol. バイオル 15, 576–587. ドイ:10.1046/j.1420-9101.2002.00429.x
CrossRef全文|Google Scholar
Carstens,B.C.,Pelletier,T.A.,Reid,N.M.,And Satler,J.D.(2013). 種の区切りで失敗する方法。 モル エコール 22, 4369–4383. 土井:10.1111/mec.12413
PubMed Abstract|CrossRef Full Text|Google Scholar
Coates,D.J.(1988). アカシア-アノマラ-コートにおける遺伝的多様性と集団遺伝子構造。 オースト J.Bot. 36:273–286. doi:10.1071/BT9880273
CrossRef全文/Google Scholar
Coates,D.J.(2000). 豊かで断片化した植物相における保全単位の定義:南西オーストラリア植物における遺伝資源と進化過程の管理への影響。 オースト J.Bot. 48, 329–339. doi:10.1071/BT99018
CrossRef全文/Google Scholar
Coates,D.J.,McArthur,S.L.,And Byrne,M.(2015). まれな風土病Banksia brownii(Proteaceae)における病原体駆動集団絶滅後の重要な遺伝的多様性の損失。 バイオル コンサーヴ 192, 353–360. doi:10.1016/j.biocon.2015.10.013
CrossRef全文|Google Scholar
Colgan,D.J.,and Paxton,J.R.(1997). 生化学的遺伝学と一般的なジェムフィッシュ、Rexea solandri(Scombroidea:Gempylidae)の西洋株の認識。、オーストラリアで。 マー… フレッシュウ… Res.48,103-118. doi:10.1071/MF96048
CrossRef全文/Google Scholar
生物多様性に関する条約(2007)。 オンラインで入手可能:https://www.cbd.int/doc/meetings/cop-bureau/cop-bur-2007/cop-bur-2007-10-14-en.pdf
Cook、C.N.、およびSgrò、C.M.(2017)。 進化的に啓発された保全を達成するために科学と政策を整列させる。 コンサーヴ バイオル 31, 501–512. 土井:10.1111/cobi.12863
PubMed Abstract|CrossRef Full Text|Google Scholar
Cook,L.G.,Edwards,R.D.,Crisp,M.D.,And Hardy,N.B.(2010). 新しい種の記述には形態学が常に必要ですか? 反転します。 シスオ… 24, 322–332. doi:10.1071/IS10011
CrossRef全文/Google Scholar
Cracraft,J.(1983). 種の概念と種分化の分析。 カー オルニトール 1, 159–187. 土井:10.1007/978-1-4615-6781-3_6
CrossRef全文|Google Scholar
Crandall,K.A.,Bininda-Emonds,O.R.,Mace,G.M.,And Wayne,R.K.(2000). 保全生物学における進化過程を考える。 トレンド-エコール… エヴォル 15, 290–295. doi:10.1016/S0169-5347(00)01876-0
Pubmed Abstract|CrossRef Full Text/Google Scholar
Cutter,A.D.,And Payseur,B.A.(2013). リンクされたサイトでの選択のゲノムシグネチャ:種間の格差を統一する。 ナット ジュネット牧師 14, 262–274. 土井:10.1038/nrg3425
PubMed Abstract|CrossRef Full Text|Google Scholar
Dayrat,B.(2005). 統合的な分類学に向けて。 バイオル J.Linn. Soc. 85, 407–415. ドイ:10.1111/j.1095-8312.2005.00503.x5
CrossRef全文|Google Scholar
de Queiroz,K.(1998). 「種の一般的な系統概念、種の基準、および種分化のプロセス」、形態:種および種分化、D.J.Howard、r S.H.Berlocher(ニューヨーク、NY:Oxford University Press)、57-75。
Google Scholar
de Queiroz,K.(2007). 種の概念と種の区切り。 シスト バイオル 56, 879–886. doi:10.1080/10635150701701083
PubMed Abstract|CrossRef Full Text|Google Scholar
Degnan,J.H.,And Rosenberg,N.A.(2009). 遺伝子ツリーの不一致、系統発生推論と多種合体。 トレンド-エコール… エヴォル 24, 332–340. 土井:10.1016/j。2009.01.009
Pubmed Abstract|CrossRef Full Text|Google Scholar
Devey,D.S.,Bateman,R.M.,Fay,M.F.,And Hawkins,J.A.(2008). 友人や親戚? 論争の的になっているヨーロッパのラン属Ophrysの系統学と種の境界。 アン ボット 101, 385–402. doi:10.1093/aob/mcm299
Pubmed Abstract|CrossRef Full Text/Google Scholar
Dizon,A.E.,Lockyer,C,Perrin,W.F.,Demaster,D.P.,Sisson,J.(1992). 株の概念を再考する-系統地理学的アプローチ。 コンサーヴ バイオル 6, 24–36. doi:10.1046/j.1523-1739.1992.610024.x
CrossRef全文/Google Scholar
Doughty,P.,Bourke,G.,Tedeschi,L.,Pratt,R.,Oliver,P.,Palmer,r.,et al. (2018). ゲヒラナナ(学名:”Gekkonidae”)は、スズメ目ゲヒラナナ科に分類される鳥類の一種である。 複合体:オーストラリアのモンスーン熱帯地方における不可解で発散的な形態学的進化、四つの新しい種の記述。 Zootaxa,4403:201-244. ドイ:10.11646/zootaxa.4403.2.1
Pubmed Abstract|CrossRef Full Text|Google Scholar
Dynesius,M.,And Jansson,R.(2014). 種内系統の持続性:種分化率の無視された制御。 進化68,923-934. 土井:10.1111/evo.12316
PubMed Abstract|CrossRef全文|Google Scholar
Edwards,S.V. とができるようになることを期待しています。 網状分裂、発散、および系統地理学-系統学continuumc。 プロク… ナトル アカド サイ… 113,8025-8032. ドイ:10.1073/pnas.1601066113
CrossRef全文|Google Scholar
Faith,D.P.(1992). 保全評価と系統発生の多様性。 バイオル 短所。 61, 1–10. 土井: 10.1016/0006-3207(92)91201-3
CrossRef全文|Google Scholar
Finston,T.L.,Johnson,M.S.,Humphreys,W.F.,Eberhard,S.M.,And Halse,S.A.(2007). 二つの広範な地下両足属の不可解な種分化は、古代の風景の歴史的な排水パターンを反映しています。 モル エコール 16, 355–365. doi:10.1111/j.1365-294X.2006.03123.x
Pubmed Abstract|CrossRef Full Text|Google Scholar
Firestone,K.B.,Elphinstone,M.S.,Sherwin,W.B.,And Houlden,B.A.(1999). 絶滅の危機に瀕している肉食性有袋類であるtiger quolls Dasyurus maculatus(Dasyuridae:Marsupalia)の系統地理学的個体群構造。 モル エコール 8, 1613–1625. doi:10.1046/j.1365-294x.1999.00745。x
CrossRef全文|Google Scholar
Fißer,C.,Robinson,C.T.,Malard,F.(2018). 種の概念のパラダイムシフトへの窓としての不可解な種。 モル エコール 27, 613–635. 土井:10.1111/mec.14486
PubMed Abstract|CrossRef Full Text|Google Scholar
Frankham,R.(2010). 生物学的保全への遺伝的アプローチの課題と機会。 バイオル コンサーヴ 143,1919–1927. doi:10.1016/j.biocon.2010.05.011
CrossRef全文|Google Scholar
Frankham,R.,Ballou,J.D.,Dudash,M.R. ら、eldridge,M. (2012). 生物多様性を保全するための異なる種の概念の意味。 バイオル コンサーヴ 153, 25–31. doi:10.1016/j.biocon.2012.04.034
CrossRef全文|Google Scholar
Fraser,D.J.,And Bernatchez,L.(2001). 適応進化的保存:保存単位を定義するための統一された概念に向かって。 モル エコール 10, 2741–2752. doi:10.1046/j.1365-294X.2001.t01-1-01411.x
PubMed Abstract|CrossRef Full Text|Google Scholar
Funk,W.C.,Mckay,J.K.,Hohenlohe,P.A. ることを示した。 保全ユニットを描写するためのゲノミクスを利用する。 トレンド-エコール… エヴォル 27, 489–496. 土井:10.1016/j。2012.05.012
PubMed Abstract|CrossRef Full Text|Google Scholar
Garnett,S.T.,And Christidis,L.(2017). 分類学アナーキーは保全を妨げる。 自然546、25-27。 doi:10.1038/546025a
PubMed Abstract|CrossRef Full Text|Google Scholar
Gippoliti,S.,Cotterill,F.P.D.,Zinner,D.,And Groves,C.P.(2018). アフリカの有蹄多様性の保全のための分類学的慣性の影響: 概要です。 バイオル 93巻、115-130頁。 ドイ:10.1111/brv。12335
PubMed Abstract|CrossRef Full Text|Google Scholar
Grant,V.(1981). 植物の種分化。 ニューヨーク、ニューヨーク:コロンビア大学プレス。PubMed Abstract/Google Scholar
Hamilton,C.W.,And Reichard,S.H.(1992). 野生植物の分類における亜種、品種および形態の使用における現在の実践。 タクソン41,485-498. doi:10.2307/1222819
CrossRef全文|Google Scholar
Harvey,M.S.,Rix,M.G.,Framenau,V.W.,Hamilton,Z.R. Johnson,M.S.,Teale,R.J.,et al. (2011). 無実の保護:短距離風土病の分類群を研究することは、保全の成果を高めます。 反転します。 シスト 25, 1–10. doi:10.1071/IS11011
CrossRefのフルテキスト|Google Scholar
こんにちは、Waples,R.S.,アーノルドシステムButlin,R.K.、ハリソン,R.G.(2003)を参照のこと。 生物学と保全における種の不確実性を理解し、直面する。 トレンド-エコール… エヴォル 18, 597–603. 土井:10.1016/j。2003.08.014
CrossRef全文|Google Scholar
Hoban,S.M.,Hauffe,H.C.,Perez-Espona,S.,Arntzen,J. W.,Bertorelle,G.,Bryja,J.,et al. (2013). 遺伝的多様性を保全政策と管理の最前線にもたらす。 コンサーヴ ジュネット リソー 5, 593–598. ドイ:10.1007/s12686-013-9859-y
CrossRef全文|Google Scholar
Isaac,N.J.,Mallet,J.,And Mace,G.M.(2004). 分類学的インフレーション:マクロ生態学と保全への影響。 トレンド-エコール… エヴォル 19, 464–469. 土井:10.1016/j。2004.06.004
PubMed Abstract|CrossRef Full Text|Google Scholar
Isaac,N.J.,Turvey,S.T.,Collen,b.,Waterman,C.,And Baillie,J. E.(2007年)。 エッジ上の哺乳類:脅威と系統発生に基づく保全の優先順位。 PLoS ONE2:e296. doi:10.1371/ジャーナル。ポネ0000296
PubMed Abstract|CrossRef Full Text|Google Scholar
Jones,M.R.,And Good,J.M.(2016). 進化的および生態学的ゲノミクスにおける標的捕獲。 モル エコール 25, 185–202. 土井:10.1111/mec.13304
PubMed Abstract|CrossRef Full Text|Google Scholar
Jorger,K.M.,And Schrödl,M.(2013). 不可解な種を記述するには? 分子分類学の実用的な課題。 フロント ゾール 10:59. 土井:10.1186/1742-9994-10-59
PubMed Abstract|CrossRef Full Text|Google Scholar
Laikre,L.,Allendorf,F.,Aroner,L.C.,Baker,S.,Gregovich,D.P.,Hansen,M.M.,et al. (2010). 生物多様性に関する条約の実施における遺伝的多様性の無視。 コンサーヴ バイオル 24, 86–88. ドイ:10.1111/j.1523-1739.2009.01425.x
Pubmed Abstract|CrossRef Full Text|Google Scholar
Laikre,L.,Palm,S.,And Ryman,N.(2005). 魚の遺伝的個体群構造:沿岸域管理への影響。 34,111–119 土井:10.1579/0044-7447-34.2.111
Pubmed Abstract|CrossRef Full Text|Google Scholar
Laity,T.,Laffan,S.W.,González-Orozco,C.E.,Faith,D.P.,Rosauer,D.F.,Byrne,M.,et al. (2015). 保全政策を知らせるためにPhylodiversity:オーストラリアの例。 サイ… 総Environ。 534, 131–143. ドイ:10.1016/j.scitotenv.2015.04.113
Pubmed Abstract|CrossRef Full Text|Google Scholar
Leache,A.D.,And Oaks,J.R.(2017). 一塩基多型(SNP)の有用性。 系統発生学のデータ。 アン エコールレヴュー シスト 48, 69–84. 土井:10.1146/annurev-ecolsys-110316-022645
CrossRef Full Text | Google Scholar
Llorens, T. M., Macdonald, B., McArthur, S., Coates, D. J., and Byrne, M. (2015). Disjunct, highly divergent genetic lineages within two rare Eremophila (Scrophulariaceae: Myoporeae). species in a biodiversity hotspot: implications for taxonomy and conservation. Biol. J. Linn. Soc. 177, 96–111. doi: 10.1111/boj.12228
CrossRef Full Text | Google Scholar
Love Stowell, S. M., Pinzone, C. A., and Martin, A. P. (2017). 遺伝的多様性のための積極的な介入への障壁を克服する。 バイオダイバーズ コンサーヴ 26, 1753–1765. 土井:10.1007/10531-017-1330-z
CrossRef全文|Google Scholar
Mace,G.M.(2004). 種の保全における分類学の役割。 フィロス トランス R.Soc. B:Biol. サイ… 359, 711–719. ドイ:10.1098/rstb。2003.1454
PubMed Abstract|CrossRef Full Text|Google Scholar
Mallet,J.(2007). ハイブリッド種分化。 自然446,279-283. 土井:10.1038/nature05706
PubMed Abstract|CrossRef Full Text|Google Scholar
Mayr,E.(1942). 系統学と種の起源。 ニューヨーク、ニューヨーク:コロンビア大学プレス。
Google Scholar
Mee,J.A.,Bernatchez,L.,Reist,J.D.,Rogers,S.M.,And Taylor,E.B.(2015). 種内保全の優先順位付けのための設計可能なユニットの特定:湖ホワイトフィッシュ種複合体に適用される階層的なアプローチ(Coregonus spp.). エヴォル Appl. 8, 423–441. 土井:10.1111/エヴァ.12247
PubMed Abstract | CrossRef Full Text | Google Scholar
Millar, M. A., and Byrne, M. (2013). Cryptic divergent lineages of Pultenaea pauciflora M.B. Scott (Fabaceae: Mirbelieae) exhibit different evolutionary history. Biol. J. Linn. Soc. 108, 871–881. doi: 10.1111/bij.12002
CrossRef Full Text | Google Scholar
Moritz, C. (1994). Defining evolutionarily significant units for conservation. Trends Ecol. Evol. 9, 373–375. doi: 10.1016/0169-5347(94)90057-4
CrossRef Full Text | PubMed Abstract
Moritz, C. (1999). 保存単位と転座:進化過程を保存するための戦略。 130件中217-228件を表示しています。 ドイ:10.1111/j.1601-5223.1999.00217.x
CrossRef全文|Google Scholar
Moritz,C.(2002). 生物多様性とそれを維持する進化過程を保護するための戦略。 シスト バイオル 51, 238–254. doi:10.1080/10635150252899752
PubMed Abstract|CrossRef Full Text|Google Scholar
Moritz,C.,And Potter,S.(2013). 保全政策計画における進化的視点の重要性。 モル エコール 22, 5969–5971. 土井:10.1111/mec.12565
PubMed Abstract|CrossRef Full Text|Google Scholar
Moritz,C.,Pratt,R.C.,Bank,S.,Bourke,G.,Bragg,J.G.,Doughty,P.,et al. (2018). オーストラリアのトカゲ(Gehyra)の種複合体における不可解な系統の多様性、体の大きさの発散、および同所性。 進化論27,54-66. 土井:10.1111/evo.13380
CrossRef全文|Google Scholar
Morrison,W.R.,Lohr,J.L.,Duchen,P.,Wilches,R.,Trujillo,D.,Mair,M.,et al. (2009). 分類学的変化が保全に及ぼす影響: それは殺すのですか、それは救うことができますか、それとも無関係ですか? バイオル 短所。 142, 3201–3206. doi:10.1016/j.biocon.2009.07.019
CrossRef全文|Google Scholar
Murphy,N.P.,Adams,M.,Guzik,M.T.,And Austin,A.D.(2013). オーストラリアの砂漠の春の両生類の異常なマイクロ風土病。 モル フィロゲネット エヴォル 66, 645–653. doi:10.1016/j.ympev。2012.10.013
Pubmed Abstract|CrossRef Full Text|Google Scholar
Murphy,N.P.,King,R.A.,And Delean,S.(2015). 種、Esuまたは集団? オーストラリアの砂漠の春の両生類の形態学的に不可解な多様性を区切ると記述する。 反転します。 シスト 29, 457–467. doi:10.1071/IS14036
CrossRef全文/Google Scholar
Nater,A.,Mattle-Greminger,M.P.,Nurcahyo,A.,Nowak,M.G.,de Manual,M.,Desai,T.,et al. (2017). 新しいオランウータン種の形態測定、行動、およびゲノムの証拠。 カー セルバイオール 27, 3487–3498. 土井:10.1016/j.cub.2017.09.047
CrossRef全文|Google Scholar
Nicotra,A.B.,Chong,C.,Bragg,J.G.,Ong,C.R.,Aitken,N.C.,Chuah,A.、ら。 (2016). オーストラリアのペラルゴニウムにおけるgenotyping-by-sequencingを介した個体群と系統学的分解。 モル エコール 25, 2000–2014. 土井:10.1111/mec.13584
PubMed Abstract|CrossRef Full Text|Google Scholar
Nistelberger,H.,Byrne,M.,Coates,D.,And Roberts,J.D.(2014a). ヤスデの強い系統地理学的構造は,半乾燥オーストラリアの帯状鉄層上の個体群間の更新世の移行を示している。 PLoS ONE9:e0093038. doi:10.1371/ジャーナル。ポネ0093038
PubMed Abstract|CrossRef Full Text/Google Scholar
Nistelberger,H.,Gibson,N.,Macdonald,B.,Tapper,S.-L.,And Byrne,M.(2014b). 生物多様性のホットスポットにおける二つのmesicレフュージアの系統地理学的証拠。 113件中454-463件を表示しています。 ドイ:10.1038/hdy.2014.46
Pubmed Abstract|CrossRef Full Text|Google Scholar
Ogilvie,H.A.,Heled,J.,Xie,D.,And Drummond,A.J.(2016). Beastの計算性能と統計的精度、および他の方法との比較。 シスト バイオル 65, 381–396. 土井:10.1093/sysbio/syv118
PubMed Abstract|CrossRef Full Text|Google Scholar
Oliver,P.M.,Keogh,J.S.,And Moritx,C.(2015). 進化の多様性をカタログ化し理解するための新しいアプローチ:オーストラリアのヘルペス学からの視点。 オースト J.ゾール 62, 417–430. doi:10.1071/ZO14091
CrossRef全文/Google Scholar
Oliver,P.M.,Laver,R.J.,Demello Martins,F.,Pratt,R.C.,Hunjan,S.,And Moritz,C.(2017). 熱帯オーストラリアにおける脊椎動物の固有種と進化的避難所の新しいホットスポット。 ダイバー ディストリブ 23, 53–66. doi:10.1111/ddi。12506
CrossRef全文|Google Scholar
Oliver,P.M.,and Lee,M.S.Y.(2010). 植物および動物学のコードは、無名の新種の出版を阻止することによって生物多様性の研究を妨げている。 タクソン59,1201-1205. doi:10.2307/20773990
CrossRef全文|Google Scholar
Padial,J.,Miralles,A.,De la Riva,I.,And Vences,M.(2010). 分類学の統合的な未来。 フロント ゾール 7:16. 土井:10.1186/1742-9994-7-16
PubMed Abstract|CrossRef Full Text|Google Scholar
Palsbøll,P.J.,Bérubé,M.,And Allendorf,F.W.(2007). 集団遺伝データを用いた管理単位の同定。 トレンド-エコール… エヴォル 22, 11–16. 土井:10.1016/j。2006.09.003
Pubmed Abstract|CrossRef Full Text|Google Scholar
Patton,J.L.,And Smith,M.F.(1994). Pocket Gophers(属Thomomys)におけるParaphyly、polyphyly、および種の境界の性質。 シスト バイオル 43, 11–26. ドイ:10.1093/sysbio/43.1.11
CrossRef全文|Google Scholar
Pierson,J.C.,Coates,D.J.,Oostermeijer,J.G.B.,Beissinger,S.R.,Bragg,J.G.,Sunnucks,P.,et al. (2016). 三大陸における絶滅危惧種の回復計画における遺伝的要因。 フロント エコール エンビロン 14, 433–440. 土井:10.1002/料金.1323
CrossRef全文|Google Scholar
Potter,S.,Bragg,J.G.,Peter,B.M.,Bi,K.,And Moritz,C.(2016). 先端の系統発生学:熱帯トカゲ、Carlia amaxにおける系統とその人口統計学的歴史を推論する。 モル エコール 25, 1367–1380. 土井:10.1111/13546
PubMed Abstract|CrossRef Full Text|Google Scholar
Potter,S.,Xue,A.T.,Bragg,J.G.,Rosauer,D.F.,Roycroft,E.J.,And Moritz,C.(2018). 更新世の気候の変化は、熱帯サバンナ全体の多様化を駆動します。 モル エコール 27, 520–532. 土井:10.1111/mec.14441/full
PubMed Abstract|CrossRef Full Text|Google Scholar
Rabosky,D.L.,Hutchinson,M.N.,Donnellan,S.C.,Talaba,A.L.,And Lovette,I.J.(2014). オーストラリアのスキンク(Scincidae:Ctenotus)の広範なグループにおける種の境界の系統発生分解。 モル フィロゲネット エヴォル 77, 71–82. doi:10.1016/j.ympev。2014.03.026
Pubmed Abstract|CrossRef Full Text|Google Scholar
Ralls,K.,Ballou,J.D.,Dudash,M.R.,Eldridge,M.D.B.,Fenster,C.B.,Lacy,R.C.,et al. (2018). 断片化された集団の遺伝的管理におけるパラダイムシフトを求める。 コンサーヴ レット 11, 1–6. 土井:10.1111/conl.124126
CrossRef全文|Google Scholar
Rannala,B.(2015). 種の境界の芸術と科学。 カー ゾール 61, 846–853. doi:10.1093/czoolo/61.5.846
CrossRef全文/Google Scholar
Riedel,A.,Sagata,K.,Suhardjono,Y.R.,Tänzler,R.,And Balke,M.(2013). ファーストトラック上の統合的な分類-生物多様性の研究におけるより多くの持続可能性に向けて。 フロント ゾール 10:15. doi:10.1186/1742-9994-10-15
PubMed Abstract|CrossRef Full Text|Google Scholar
Rieseberg,L.H.,And Willis,J.H.(2007). 植物の種分化。 科学317,910-914. 土井:10.1126/科学。1137729
PubMed Abstract|CrossRef Full Text|Google Scholar
Rosauer,D.F.,Blom,M.P.K.,Bourke,G.,Catalano,S.,Donnellan,S.,Mulder,E.,et al. (2016). 系統地理学、ホットスポットと保全の優先順位:オーストラリアの上端からの例。 バイオル 短所。 204, 83–93. doi:10.1016/j.biocon.2016.002
CrossRef全文|Google Scholar
Rosauer,D.F.,Byrne,M.,Blom,M.P.K.,Coates,D.J.,Donnellan,S.,Doughty,P.,et al. (2018). オーストラリアのキンバリーにおける進化的多様性のための現実世界の保全計画は、不確実な分類学Conservを回避する。 レット 1112438. 土井:10.1111/conl.12438
CrossRef全文|Google Scholar
Rosenblum,E.B.,Sarver,B.A.J.,Brown,J.W.,Des Roches,S.,Hardwick,K.M.,Hether,T.D.,et al. (2012). ゴルディロックスはサンタロザリアを満たしています:一時的な種分化モデルは、時間スケール全体の多様化のパターンを説明しています。 エヴォル バイオル 39, 255–261. 土井:10.1007/11692-012-9171-x
Pubmed Abstract|CrossRef Full Text|Google Scholar
Roux,C.,Fraïsse,C.,Romiguier,J.,Anciaux,Y.,Galtier,N.,Bierne,N.(2016). ゲノムの発散の連続に沿って種分化のグレーゾーンに光を流します。 プロスバイオール 14:e2000234. doi:10.1371/ジャーナル。プビオ2000234
PubMed Abstract|CrossRef Full Text|Google Scholar
Ryder,O.A.(1986). 種の保全と系統学–亜種のジレンマ。 トレンド-エコール… エヴォル 1, 9–10. 土井:10.1016/0169-5347(86)90059-5
CrossRef全文/Google Scholar
Sampson,J.F.,Byrne,M.,Yates,C.J.,Gibson,N.,Thavornkanlapachai,R.,Stankowski,S.,et al. (2014). 現代の花粉媒介遺伝子移民は、断片化された風景の中でまれな低木の歴史的な分離を反映しています。 112件中172-181件を表示しています。 ドイ:10.1038/hdy.2013.89
PubMed Abstract|CrossRef Full Text|Google Scholar
Santamaría,L.,And Méndez,P.F.(2012). 生物多様性政策における進化:現在のギャップと将来のニーズ。 エヴォル Appl. 5, 202–218. 土井:10.1111/j.1752-4571.2011.00229.x
Pubmed Abstract/CrossRef Full Text/Google Scholar
Schlick-Steiner,B.C.,Steiner,F.M.,Seifert,B.,Stauffer,C.,Christian,E.,And Crozier,R.H.(2010). 統合分類学:生物多様性を探索するためのマルチソースアプローチ。 アンヌ エントモル牧師。 55, 421–438. doi:10.1146/annurev-ento-112408-085432
PubMed Abstract|CrossRef Full Text/Google Scholar
Seehausen,O,Takimoto,G.,Roy,D.,And Jokela,J.(2008). 変化する環境におけるハイブリダイゼーションを伴う種分化の逆転と生物多様性のダイナミクス。 モル-エコール 17, 30–44. doi:10.1111/j.1365-294X.2007.03529。x
Pubmed Abstract|CrossRef Full Text|Google Scholar
Shepherd,K.A.,Thiele,K.R.,Sampson,J.,Coates,D.J.,And Byrne,M.(2015). 乾燥した西オーストラリア州の二つの遺伝的に異なるが、形態学的に不可解な集団を含むAtriplex(Chenopodiaceae)のまれな、新しい種:分類学と保全への影響。 オースト シスト ボット 28, 234–245. 土井:10.1071/SB15029
CrossRef全文/Google Scholar
Singhal,S.,And Bi,K.(2017). 歴史は混乱をきれいにする:熱帯縫合線の地帯の発散そしてintrogressionを運転することの時間の影響。 進化71、1888年-1899年。 土井:10.1111/evo.13278
PubMed Abstract|CrossRef Full Text|Google Scholar
Singhal,S.,Hoskin,C.,Couper,P.,Potter,S.,Moritz,C.(2018). 謎の種を解決するためのフレームワーク:オーストラリアの湿った熱帯地方のトカゲからのケーススタディ。 シスト バイオル . 土井:10.1093/sysbio/syy
Pubmed Abstract|CrossRef Full Text|Google Scholar
Singhal,S.,And Moritz,C.(2013). 系統学的系統間の生殖分離は、発散を伴うスケールである。 プロク… ロイ Soc B280:20132246. ドイ:10.1098/rspb.2013.2246026
Pubmed Abstract|CrossRef Full Text|Google Scholar
Steane,D.A.,Potts,B.M.,McLean,E.,Collins,L.,Prober,S.M.,Stock,W.D.,et al. (2015). ゲノムワイドスキャンは、乾燥適応ユーカリの不可解な集団構造を明らかにする。 ツリー-ジュネット 11時33分現在、の最新リンクです。 土井:10.1007/11295-015-0864-z
CrossRef全文|Google Scholar
Struck,T.H.,Feder,J.L.,Bendiksby,M.,Birkeland,S.,Cerca,J.,Gusarov,V.I.,et al. (2017). 謎の種に隠された進化過程を見つける。 トレンド-エコール… エヴォル 33, 153–163. 土井:10.1016/j。2017.11.007
Pubmed Abstract|CrossRef Full Text|Google Scholar
Sukumaran,J.,And Knowles,L.L.(2017). 多種合体は種ではなく構造を区別する。 プロク… ナット アカ… サイ… アメリカ114、1607-1612。 ドイ:10.1073/pnas.1607921114
PubMed Abstract|CrossRef Full Text|Google Scholar
Swarts,N.D.,Clements,M.A.,Bower,C.C.,And Miller,J.T.(2014). 形態学的に類似した、性的に欺瞞的な、非常に絶滅の危機に瀕している蘭の複合体における保全単位を定義する。 バイオル 短所。 174, 55–64. doi:10.1016/j.biocon.2014.03.017
CrossRef全文|Google Scholar
Tautz,D.,Arctander,P.,Minelli,A.,Thomas,R.H.,And Vogler,A.P.(2003). DNA分類学のための嘆願。 トレンド-エコール… エヴォル 18, 70–74. 土井:10.1016/S0169-5347(02)00041-1
CrossRef全文/Google Scholar
Taylor,H.R.,Dussex,N.,and van Heezik,Y.(2017). 保全の実践者のための実装への障壁を特定することにより、保全遺伝学のギャップを埋める。 グロブ エコール コンサーヴ 10, 231–242. 土井:10.1016/j.gecco.2017.04.001
CrossRef全文|Google Scholar
Thomson,S.A.,Pyle,R.L.,Ahyong,S.T.,Alonso-Zarazaga,M.,Ammirati,J.,Araya,J.F.,et al. (2018). 地球規模の保全には科学に基づく分類が必要である。 プロスバイオール 16:e2005075. 土井:10.1371/日記プビオ2005075
PubMed Abstract|CrossRef Full Text|Google Scholar
Vieites,D.R.,Wollenberg,K.C.,Andreonec,F.,Köhler,J.,Glaw,F.,and Vences,M.(2009). 統合的な両生類の目録によって証明されるマダガスカルの生物多様性の広大な過小評価。 プロク… ナトル アカド サイ… 米国106、8267-8272。 ドイ:10.1073/pnas.0810821106
CrossRef全文
Vogler,A.P.,and de Salle,R.(1994). 保全管理の単位を診断します。 短所。 バイオル 8, 354–363. doi:10.1046/j.1523-1739.1994.08020354.x
CrossRef全文|Google Scholar
Waples,R.S.(1991). 太平洋サーモン、Oncorhynchus spp。、および絶滅危惧種法の下で”種”の定義。 マー… フィッシュ 昭和53年11月22日
Google Scholar
Waples,R.S.(1995). “進化的に重要な単位と絶滅危惧種法の下での生物多様性の保全,”進化と水生生態系:人口保全におけるユニークな単位の定義,編J.L.ニールセン(Bethesda,MD:American Fisheries Society),8-27.
Google Scholar
Waples,R.S.,Gustafson,R.G.,Weitkamp,L.A.,Myers,J.M.,Jjonson,O.W.,Busby,P.J.,et al. (2001). 太平洋岸北西部からのサケの多様性を特徴付ける。 J.Fish Biol. 59, 1–41. ドイ:10.1111/j.1095-8649.2001.01376x
CrossRef全文/Google Scholar
Waples,R.S.,Nammack,M.,Cochrane,J.F.,and Hutchings,J.A.(2013). 二つの行為の物語:カナダと米国での絶滅危惧種のリストの慣行。 バイオサイエンス63,723-734. ドイ:10.1093/バイオサイエンス/63.9.723
CrossRef全文|Google Scholar
Waples,R.S.,Punt,A.E.,And Cope,J.M.(2008). 海洋資源の管理に遺伝子データを統合する:どのように我々はそれをより良くすることができますか? 魚9、423-449。 ドイ:10.1111/j.1467-2979.2008.00303.x
CrossRef全文/Google Scholar
Wege,J.A.,Thiele,K.R.,Shepherd,K.A.,Butcher,R.,Macfarlane,T.D.,And Coates,D.J.(2015). 生物多様性の景観における戦略的分類:希少であまり知られていない植物相の保全成果を最大化するための新しいアプローチ。 バイオダイバーズ コンサーヴ 24, 17–32. 土井:10.1007/10531-014-0785-4
CrossRef全文/Google Scholar
Whitlock,R.(2014). 適応性と中立性の遺伝的多様性と生態学的構造と機能の関係:メタ分析。 J.Ecol. 102, 857–872. doi:10.1111/1365-2745.12240
PubMed Abstract|CrossRef Full Text|Google Scholar
Wiley,E.O.(1978). 進化種の概念は再考された。 シスト バイオル 27, 17–26. doi:10.2307/2412809
CrossRef全文|Google Scholar
Wilson,E.O.(2017). 生物多様性の研究は、地面に多くのブーツを必要とします。 ナット エコール エヴォル 1, 1590–1591. 土井:10.1038/s41559-017-0360-p>
PubMed Abstract|CrossRef全文|Google Scholar
Yang,Z.H.,And Rannala,B.(2014). 複数の遺伝子座からのDNA配列データを使用して誘導されていない種の区切り。 モル バイオル エヴォル 31, 3125–3135. doi:10.1093/molbev/msu279
Pubmed Abstract|CrossRef Full Text/Google Scholar
Yeates,D.K.,Seago,A.,Nelson,L.,Cameron,S.L.,Joseph,L.,And Trueman,W.H.(2011). 統合的な分類、または反復的な分類学? シスオ… Ent. 36, 209–217. doi: 10.1111/j.1365-3113.2010.00558.x
CrossRef Full Text | Google Scholar