az 1960-as évek elején a biológusok elkezdték vizsgálni, hogy a fehérjék hogyan különböző fajokban fejlődik a szekvencia szintjén (Zuckerkandl 1962, margoliash 1963, Doolittle& blombäck 1964). A vizsgált fehérjék közé tartozott a hemoglobin (Zuckerkandl & Pauling 1962), a citokróm C (Margoliash 1963) és a fibrinopeptidek (Doolittle & Blombäck 1964). Ezek a korai vizsgálatok figyelemre méltó felfedezéshez vezettek: úgy tűnt, hogy a különböző fajok fehérjeszekvenciái közötti különbségek száma nagyjából arányos volt a fajok divergenciája óta eltelt idővel (1.ábra).
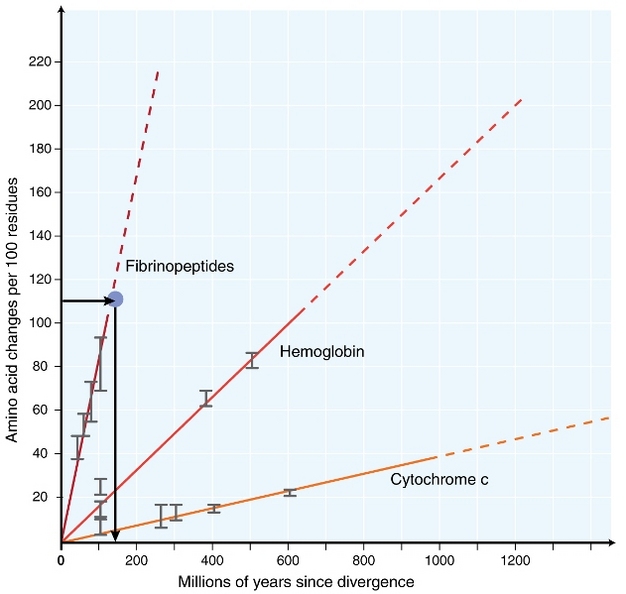
Zuckerkandl& Pauling (1965) hasonlította össze az aminosavpótlások állandó időbeli felhalmozódását az órák rendszeres “kullancsaival”, és kijelentette, hogy “létezik molekuláris evolúciós óra”. Így a “molekuláris óra” kifejezést eredetileg úgy alkották meg, hogy leírják az aminosavak változásait, amelyek a fajok eltérése óta arányosan fordulnak elő.
az első használat óta a “molekuláris óra” kifejezést számos különböző kontextusban használják. Manapság gyakran használják egyszerűen a DNS vagy a fehérjék szekvenciájában felhalmozódott változások vagy “helyettesítések” számára. Az egy meghatározott időegységre jutó helyettesítések számát A molekuláris óra “sebességének” lehet leírni, amely ebben az összefüggésben egyenértékű az “evolúciós sebességgel”. Ne feledje, hogy a kezdeti konnotáció bizonyos esetekben továbbra is fennáll. Például, amikor az emberek azt állítják, hogy “molekuláris órát feltételeztünk” (különösen filogenetikai elemzésekben), ez azt jelenti, hogy a helyettesítések számát az idő múlásával folyamatosan feltételezték.
az állandó molekuláris óra fogalma rendkívüli következményekkel jár az evolúciós biológusok számára. Ha az eredetileg javasolt állandó molekuláris óra valóban létezne, az evolúciós események időzítésének következtetése meglehetősen egyszerű problémává válna (1.ábra). Ugyanakkor világossá vált, hogy a helyettesítések nem fordulnak elő folyamatosan az idő múlásával különböző sorokban (Kumar 2005). Mindazonáltal a molekuláris óra fogalma rendkívül befolyásos volt a molekuláris evolúció területén. A molekuláris óra koncepciója által inspirált egyik legfontosabb ötlet a molekuláris evolúció semleges elmélete.
A molekuláris óra és semlegesség
amikor egy állandó molekuláris óra gondolata először merült fel, azt hitték, hogy az aminosav vagy nukleotid szubsztitúciók alapjául szolgáló domináns evolúciós erő a természetes szelekció. Ezt a gondolkodásmódot követve egy állandó molekuláris óra azt jelzi, hogy a különböző fajok adaptív szubsztitúciói az idő múlásával folyamatosan fordulnak elő. Nehéz azonban megmagyarázni, hogy az adaptív helyettesítések hogyan fordulnak elő ilyen óraszerű módon. Elméletileg az adaptív mutációk sorsát több evolúciós paraméter határozza meg, mint például a mutáció szelektív előnyének erőssége, a hatékony populáció mérete és az adaptív mutációs arányok (Kimura 1983). Ezek a paraméterek valószínűleg fajonként, sőt fajon belül is különböznek, a specifikus mutációktól és a környezetekkel való kölcsönhatásuktól függően.
ehelyett Kimura (1968, 1969) azt javasolta, hogy a legtöbb molekuláris szintű változásnak kevés funkcionális következménye van, vagy “semleges”. Ha egy mutációnak nincs fitnesz következménye, sorsát a populációban teljesen véletlenszerű véletlen határozza meg. Ez azt jelenti, hogy nem tudjuk megjósolni, hogy egy adott semleges mutáció végül rögzül-e a populációban. Azonban a populációban a semleges helyettesítések aránya megjósolható, mivel egyetlen paramétertől, nevezetesen a mutációs aránytól függ (Kimura 1968).
képzeljünk el egy n számú haploid egyedet. Ha semleges mutációk fordulnak elő u sebességgel egyénenként generációnként, az összes mutáció egy generációban n-szer u. mivel ezek az új mutációk semlegesek, sorsukat véletlenszerűen határozzák meg. Más szavakkal, minden mutációnak egyenlő esélye van a rögzítés elérésére (ami “helyettesítéshez” vezet). Annak a valószínűsége, hogy minden új semleges mutáció eléri a rögzítést, mivel helyettesítés történt, egyszerűen 1 / N. Az arány a cserék kerül kiszámításra, mint az, hogy számos új mutációk minden generáció (Nu) szorozva a valószínűsége, minden új mutáció eléri rögzítés (1/N), ami egyenlő u. Más szóval, a semleges mutációk, az arány a helyettesítő egyenlő az arány a mutáció!
ezért, ha a legtöbb mutáció semleges (a semleges elméletben javasolt), és ha a mutációs arányok az idő múlásával állandóak, a szubsztitúcióknak az idő múlásával is folyamatosan kell történniük. Ezután meg kell figyelnünk az óraszerű rendszeres helyettesítéseket molekuláris szinten. Kimura (1969)tehát a viszonylag állandó molekuláris óra megfigyelését a fehérjeszekvenciákban a molekuláris evolúció semleges elméletének erős támogatásaként tekintette.
A semleges molekuláris óra működésének vizsgálata
a semleges elmélet szerint az a kérdés, hogy a helyettesítési arányok idővel állandóak-e vagy sem, egyenértékű-e azzal, hogy a semleges mutációs arányok állandóak-e az idő múlásával. Emiatt számos későbbi tanulmány a semleges helyek adatainak elemzésére összpontosított annak meghatározására, hogy a semleges mutációs arányok valóban állandóak-e az idő múlásával. Röviden áttekintjük ezeket a tanulmányokat, mielőtt megvitatnánk a molekuláris órákkal kapcsolatos elméleti vitákat. A fehérjemolekuláris órák elemzése is folytatódott, de a fehérje molekuláris órák változásait övező viták nagyon különböznek a semleges molekuláris órákkal kapcsolatos vitáktól, és nem szerepelnek ebben a cikkben. Az érdeklődő olvasóknak konzultálniuk kell Gillespie-vel (1991), Kumarral (2005), Kim and Yi-vel (2008), valamint Bedford et al. (2008).
a semleges molekuláris órák legtöbb empirikus elemzése arra a tételre támaszkodik, hogy a semleges mutációs arányok semleges szubsztitúciós arányokból következtethetők (Kimura 1968, 1969). A gyakorlatban minden egyes vizsgálat a genom egy bizonyos típusát semleges helyként határozza meg, és összehasonlítja ezen helyek helyettesítési arányát a vonalak között.
mely helyek a genomban valóban semlegesek, nem lehet teljesen meghatározni, de a tudósok számos hasznos proxyval álltak elő. A genomszekvenálás kora előtt a legtöbb rendelkezésre álló szekvencia adat a fehérjekódoló DNS-szekvenciákból származott. A vizsgálatok gyakran a fehérjekódoló DNS-szekvenciákat két típusú helyre osztják (Wu & Li 1985). Az első típusú helyek közé tartoznak azok, amelyek bármilyen változás vezetne aminosav szubsztitúciók, vagy “nonsynonim helyek”. A második típusú helyek, “szinonim helyek”, magában foglalja azokat, amelyek kódolják a “degenerált” pozíciókat a kodon táblázatban, ahol a változás nem vezet aminosav-helyettesítéshez. Például a TCT és a TCC egyaránt kódolja a szerint. Ha ezeknek a kodonoknak a harmadik helyzete megváltozik, akkor továbbra is ugyanazokat az aminosavakat állítják elő. Az ilyen helyettesítések kevésbé láthatók a természetes szelekció számára. Következésképpen a szinonim helyeken lévő molekuláris óráknak közelebb kell lenniük a semleges molekuláris órához, mint a nem szinonim órákhoz.
ahogy a szekvenálási technikák előrehaladtak, egyes vizsgálatok intronokat használtak semleges helyek proxyiként, mivel nem épülnek be az érett mRNS-be, így nagyobb valószínűséggel semlegesek (Yi et al. 2002). Sorozatok inaktív transposable elemek egészül ki hosszú mielőtt faj divergencia is gyakran alkalmazott (ezek gyakran a továbbiakban ősi ismétli’, pl., Thomas et al., 2003). Végül néhány tanulmány nem kódoló DNS-szekvenciákat (minden szekvenciát a teljes genom-igazításokból kivont fehérje-kódoló DNS-szekvenciák eltávolítása után) használt a semleges molekuláris órák tesztelésére (Elango et al. 2006, Huttley et al. 2007).
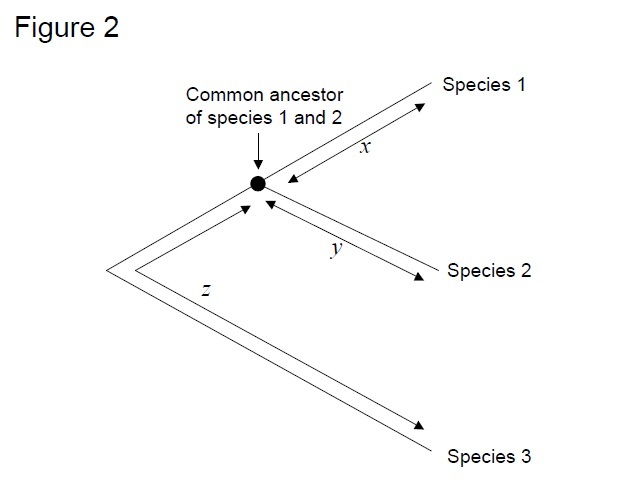
a leggyakrabban használt teszt az úgynevezett “relatív sebességteszt” (Sarich & Wilson 1973). Kezdetben a helyettesítő árak egységnyi idő alatt becsülték elosztjuk a teljes száma különbségek (szubsztitúció) között fehérjék különböző fajok által a divergencia idő, várható a fosszilis adatok (1.Ábra). A fosszilis rekordok azonban sok összehasonlításhoz nem állnak rendelkezésre, és nagy hibahatárokkal társulnak. A relatív sebesség teszt legyőzi a fosszilis nyilvántartások szükségességét (2.ábra). Mindaddig, amíg létezik egy outgroup szekvencia a két érdekes vonalhoz, meghatározhatjuk, hogy a két ág ugyanazt vagy különböző molekuláris órákat követi – e, anélkül, hogy tudnánk az eltérés abszolút idejéről (2.ábra).
melyek a semleges molekuláris órák meghatározói?
a semleges molekuláris órákkal kapcsolatos viták középpontjában álló szinte minden ellentmondás abból a kérdésből ered, hogy mi a mutációk fő forrása. Ez a kérdés közvetlenül kapcsolódik a mutáció mintáinak megértéséhez, amelyek az evolúciós változás és a genetikai betegség végső forrása. Továbbá az összehasonlító genomikában alapvető kérdés annak megértése, hogy a mutációs arányok hogyan változnak a vonalakon belül és a genomokon belül, amelynek célja a szekvencia-összehasonlítások alkalmazása a funkcionálisan fontos genomikus régiók azonosítására.
tehát mi határozza meg a semleges mutációs arányokat? A semleges molekuláris órák egyik legfontosabb hozzájárulója a generációs idők lineage-specifikus változása. A kezdetektől fogva az állandó semleges molekuláris óra gondolatát úgy tekintették, hogy ellentétes a germline mutáció molekuláris mechanizmusaival. Régóta úgy vélik, hogy a legtöbb mutáció a CSÍRAVONALAK DNS-replikációjának hibáiból ered (Haldane 1947, Muller 1954). Mivel a mutációk akkor fordulnak elő, amikor a csíravonal DNS-t a következő generációra replikálják, a generációk számának arányában kell felhalmozódniuk, nem pedig az abszolút időmennyiségben. Ezért, ha mi, mint a számok, a cserék, hogy a felhalmozódott két vonalak segítségével, mivel a divergencia, a lineage hosszabb generációs idő, hogy átesett kevesebb DNS-replikáció események, akkor harbor kevesebb cserék képest a lineage a rövidebb generációs idő. Következésképpen a molekuláris órának lassabban kell futnia a hosszabb generációs időkkel rendelkező fajokban. Ezt az elképzelést “generációs időhatásnak” nevezik.
valójában a generációs időhatást először a főemlősök tanulmányaiban figyelték meg, még a molekuláris óráról szóló vita előtt. Morris Goodman, aki immunológiai módszereket alkalmazott a fajok rokonságának vizsgálatára abban az időben, megfigyelte, hogy az egyes fehérjék eltérésének sebessége csökkent a majmokban, különösen az emberekben, összehasonlítva az óvilági majmokkal (Goodman 1961, 1962, 1963). Ezt a hatást “hominoid sebesség lassulásának” nevezik. Mivel a hominoidok hosszabb generációs időkkel rendelkeznek, mint az óvilági majmok, ez a megfigyelés a generációs időhatással magyarázható.
Wu & Li (1985) voltak az elsők, akik DNS-szekvencia-adatok felhasználásával tesztelték a generációs hatás hipotézist. 11 főemlős és rágcsáló gén adatait használták fel. Mivel a főemlősök sokkal hosszabb generációs idővel rendelkeznek, mint a rágcsálók, a molekuláris órának gyorsabbnak kell lennie rágcsálókban a főemlősökhöz képest. Valójában azt találták, hogy a szinonimák esetében a rágcsálók a főemlősökhöz képest a molekuláris evolúció sebességének körülbelül kétszeresét mutatják (Wu & Li 1985). A nonsynonim webhelyek esetében azonban ilyen hatás nem található. Más szóval, a semleges molekuláris óra, de nem az aminosav molekuláris óra, gyorsabban ketyeg a rágcsáló vonalában, mint a főemlős vonal, amely jól illeszkedik a generációs időhatáshoz.
a későbbi vizsgálatok további támogatást nyújtottak a hominoid sebesség lelassulásához (Li & Tanimura 1987, Bailey et al. 1991) és a rágcsálók és a főemlősök közötti aránykülönbség (Gu & Li 1992, Huttley et al. 2007). Ezenkívül még kisebb filogenetikai skálákban is megfigyelhető volt az aránykülönbség, különösen a főemlősöknél: például az emberi molekuláris óra lassabban fut, mint a csimpánz molekuláris órája (Elango et al. 2006); az újvilági majmok aránya gyorsabb, mint a hominoidok és az óvilági majmok aránya (Steiper & Young 2006). Az ezekben a vizsgálatokban megfigyelt molekuláris órák eltérő aránya minőségileg összhangban van a generációs időhatással.
a vonalak közötti tényleges különbségek azonban mennyiségi szempontból nem felelnek meg a generációs idők különbségének. Például Kumar & Subramanian (2002) kimutatta, hogy bár a főemlősök és a rágcsálók közötti nemzedékidők közötti különbség sokkal nagyobb, mint az emberek és az óvilági majmok között, a molekuláris órák megfigyelt különbségei hasonlóak ebben a két összehasonlításban. Érdemes megjegyezni, hogy Kumar & Subramanian (2002) speciális statisztikai szűrőket használt a “heterogén” helyettesítési mintákat mutató adatok eltávolítására, amelyek torzítást okozhattak a lassan fejlődő szekvenciák felé (Yi et al. 2002). Mindazonáltal a főemlősök és rágcsálók molekuláris óráinak különbsége sokkal kisebbnek tűnik, mint amit Wu & Li (1985) eredetileg javasolt. Például, Huttley et al. (2007) több faj teljes genomjának összehangolását elemezte, beleértve az oposszumot is, és kimutatta, hogy az eutheriai vonal és az oposszumvonal közötti sebességkülönbség (~30%) sokkal nagyobb, mint az emberi és az egér vonalai közötti sebességkülönbség (~14%). Ezek a példák azt mutatják, hogy a molekuláris órák közötti különbségek mértéke az adatkészletek és a statisztikai módszerek különbségei miatt jelentősen eltér a különböző vizsgálatok között. Azt is mutatják, hogy a vonalak közötti aránykülönbségeket nem lehet teljes mértékben elszámolni a generációs idők különbségével. Nyilvánvaló, hogy vannak más közreműködők a semleges molekuláris órákhoz.
valójában úgy tűnik, hogy a generációs időktől eltérő élettörténeti tulajdonságok a molekuláris órákkal együtt változnak. Martin & Palumbi (1993) kimutatta, hogy a DNS molekuláris órái gyorsabban futnak a kis testméretű fajokban. Ez a megfigyelés azt a hipotézist eredményezte, hogy az anyagcsere-arányok a molekuláris órák fontos meghatározói. A magas metabolikus ráta nagyszámú mutagén oxigéngyököket termel, ami növeli a mutációs arányokat (Rand 1994). Mivel a metabolikus ráta és a testméret általában együtt változik a generációs időkkel, nehéz volt megkülönböztetni, hogy ezek közül melyik a molekuláris órajel fő meghatározója. Tsantes & Steiper (2009) a főemlősök adatai alapján azt javasolta, hogy az első reprodukciókor, a testméret helyett, a molekuláris órák fő meghatározója. Mivel az életkor az első reprodukcióban tükrözi a generációs időhatást, ez a tanulmány támogatja azt az elképzelést, hogy a generációs időhatás a molekuláris óra fő meghatározója. Ez a tanulmány azonban továbbra is korlátozott számú vonalon alapul (négy fajpárt használtak). Így a testméret, a nemzedék-idő hatás és a metabolikus ráta megkülönböztetése továbbra is fontos kérdés a semleges molekuláris órák általánosításában és megértésében.
továbbá egyre inkább értékelték azoknak a tényezőknek a fontosságát, amelyek nem változnak együtt a generációs időkkel, például a DNS-metilációval. A DNS-metiláció a különböző taxonokban található genomikus DNS kémiai módosítása. Az állati genomokban a DNS-metiláció szinte kizárólag citozinokban, majd guaninokban (úgynevezett “CpG-kben”) fordul elő. A metilezett citozinok viszont kémiai instabilitás miatt gyorsan mutálódnak a timinokhoz (Bird 1980). Valójában az emberi genomban a DNS-metiláció által okozott mutációk több mint nagyságrenddel gyakrabban fordulnak elő, mint más mutációk (Nachman & Crowell 2000, Elango et al. 2008). Mivel a DNS-metiláció által okozott mutációk nagyrészt a DNS-replikációtól függetlenül fordulnak elő, az ilyen mutációk különböző molekuláris órákat követhetnek, mint mások. Pontosabban, a generációs időfüggőség helyett a DNS-metiláció által okozott mutációk egy időfüggő molekuláris órát követhetnek, amely hasonló ahhoz, amit Zuckerkandl & Pauling (Kim et al. 2006)!
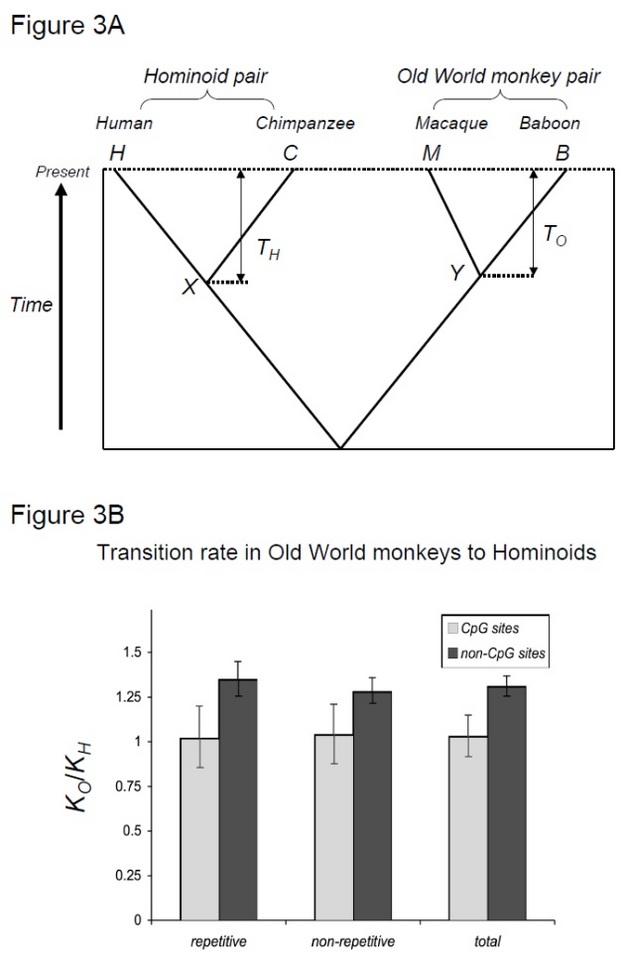
a hipotézis teszteléséhez, Kim et al. (2006) összehasonlítva az ember-csimpánz divergenciát a makákó-pávián divergenciával, két fajpárral, amelyek hasonló eltérési időkkel rendelkeznek, de különböző generációs időkkel (Steiper et al. 2004). Az ember-csimpánz párnak (a hominoid párnak) sokkal hosszabb a generációs ideje, mint a makákó-pávián párnak (az óvilági majompárnak). Ez a tanulmány kimutatta, hogy a nem CpG helyszíneken az Old World monkey pair körülbelül 30%-kal több helyettesítést halmozott fel, ami a fent említett hominoid sebesség lassulási hatásával magyarázható. Ezzel szemben a CpG-helyszíneken a molekuláris órák hasonló számú szubsztitúciót mutattak hominoid és Old World monkey párokban (3.ábra). Így az időfüggő és generációs időfüggő molekuláris órák együtt léteznek ugyanazon genomokon belül. Az a feltételezés, hogy egyetlen molekuláris óra létezhet egy adott vonalra, már nem érvényes, mert a domináns mutációs erők a genomikus régiók között változnak.
következtetések
az állandó molekuláris óra fogalmát kezdetben korlátozott mennyiségű fehérjeszekvencia adat alapján javasolták. Annak ellenére, hogy a későbbi vizsgálatok azt mutatták, hogy egy ilyen megfigyelés nem általános minta az aminosavakban, jelentős hatással volt a molekuláris evolúció területére, különösen a molekuláris evolúció semleges elméletének kifejlesztésére a DNS-szekvencia adatokra. A semleges elméletet követve a tanulmányok a semleges mutációs arányok változási mintáinak feltárására összpontosítottak. Az elmúlt néhány évtizedben megfigyeltük, hogy a molekuláris órák különböző sebességgel futnak a vonalak között. Ezenkívül a variáció mértéke a különböző típusú adatoktól és az alkalmazott konkrét statisztikai módszerektől függően változhat. A generációs időhatás továbbra is minőségi szinten tart, de nem elegendő a semleges mutációs arányok mennyiségi változásának magyarázatához a vonalak között. Élet-történelem vonások, illetve a nem-replikáció függő mutációk, mint azok által okozott DNS-metiláció is fontos támogatója genom molekuláris órák. Valójában a molekuláris órák különböző típusait még a genomban is megfigyelik, mivel a domináns mutációs bemenetek különböző genomi régiók között változnak. Így ahelyett, hogy minden genomra egyetlen semleges molekuláris órát feltételeznének, a jövőbeli tanulmányoknak arra kell törekedniük, hogy felfedezzék a genom semleges molekuláris órák variációját, hogy megismerjék a genomikus mutációs tájakat. Ez az információ nemcsak a molekuláris evolúciót és a genetikai betegséget szabályozó nyersanyag megértéséhez hasznos, hanem olyan kritikus komponens is, amely befolyásolja az Összehasonlító és funkcionális genomikai elemzéseket a funkcionális genomiális régiók azonosítása érdekében.