Bevezető
a Nemzetközi természetvédelmi politika felismeri a biológiai sokféleség három szinten, az ökoszisztéma, a faj genetikai, illetve, hogy a vezetés arra kell törekednie, hogy megtartja mind a három (a Biológiai Sokféleség Egyezmény, 2007). Ez egyértelműen tükröződik a biológiai sokféleségről szóló, 2010-ben elfogadott Egyezményben (https://www.cbd.int/sp/targets/), ahol a célokban és célokban nem csak az ökoszisztémákra és a fajokra, hanem a genetikai sokféleségre is külön hivatkozás található. Még jelenlegi megközelítések a biológiai sokféleség megőrzése vagy nagyrészt a földrajzi területeken, ökoszisztémák ökológiai közösségek, faj, kevesebb figyelmet a genetikai sokszínűség, valamint a faj-populációs folytonosság. Valójában a természetvédelmi szakemberek többsége a fajokra összpontosít, mint a biodiverzitás mutatójára, figyelembe véve az intenzíven kezelt veszélyeztetett fajok intraspecifikus egységeit. Ebben az összefüggésben természetes vágy, hogy egyszerű kategorizálásokat alkalmazzunk-elnevezett fajok, valamint más nevű taxonok (azaz alfajok), néha fajon belüli védelmi egységek. Különösen a fajok biztosítják a biológiai sokféleség közös mércéjét, amely sok tudományos és gazdálkodási törekvést támaszt alá (Mace, 2004; Wilson, 2017). A természetvédelem alapvető egységeként a fajokat gyakran használják a biológiai sokféleség értékének számszerűsítésére a védett területeken belüli Fajok jegyzékei, a veszélyeztetett fajok azonosítása a joghatóságokon belül, valamint a biológiai biztonsági eljárások alapjaként. Ezenkívül a fajok jellemzően azok az entitások, amelyekkel természetvédelmi kérdéseket kommunikálunk a nyilvánosság számára.
Ez a lendület a diszkrét egységek egyszerű azonosítására a természetben több, kapcsolódó valósággal ellentétes. Először is, a speciáció nem azonnali. Inkább ez általában egy elhúzódó folyamat (Dynesius and Jansson, 2014; Roux et al., 2016-ban), halad a elszigetelt populációk, talán témát, hogy a különböző kiválasztási nyomás, hogy egyértelmű faj hosszú idő elteltével, a gyakori feloldására által okozott környezeti változás vagy egyszerű genetikai összevonása, mint tartományok újra (1. Ábra; Seehausen et al., 2008; Rosenblum et al., 2012). Másodszor, a legismertebb fajok nem genetikailag egységesek. A divergencia (sodródás és elhúzódó izoláció) és a retikuláció (geneflow, hibridizáció, introgresszió és rekombináció) hatékonyan alakítja a fajokon belüli földrajzi és genomikai variációkat (Cutter and Payseur, 2013; Edwards et al., 2016) és egyes fajok hosszú, elszigetelt populációkba rendezhetők, amelyek közül néhány teljes, de talán morfológiailag rejtélyes fajként (Bickford et al., 2007; Jorger and Schrödl, 2013). Harmadszor, a régóta elismert, megnevezett fajok nem egyenértékűek azzal, hogy mennyi evolúciós örökséget képviselnek-egyszerűen fogalmazva, egy kacsacsőrű emlős (mint egy ősi eltérő vonal képviselője) többet ér, mint egy nemrégiben eltérített rágcsáló (Isaac et al., 2007). Negyedszer, a fajmegjelöléseket nem kőbe öntik, hanem hipotézisek, amelyeket meg kell vizsgálni, mivel új bizonyítékok állnak rendelkezésre (Hey et al., 2003). E kérdések mindegyike összetettséget és kétértelműséget teremt a fajok általános körülhatárolásában, valamint a természetvédelmi figyelmet rájuk vagy azok összetevőire fordítja. Ezenkívül a fajok elhatárolása is különböző fajfogalmaknak és diagnosztikai módszereknek van kitéve, ami nagy megdöbbenést okozott a természetvédelmi biológusok körében (Mace, 2004; Frankham et al., 2012; Garnett and Christidis, 2017).
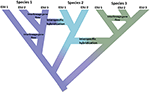
1.ábra. Az izolált populációktól az egyértelmű fajokig terjedő folytonosság, amelyet a színváltozás mutat a különböző fajokat képviselő színekkel. Néhány vonalak segítségével felmerülő miatt lakosság elszigeteltség alakulhat igaz faj, a belső csomópontok a fa képviselő kezdeményezése a speciation folyamat. Más vonalak nem alakulnak ki valódi fajokká, de idővel eltérnek egymástól, jelezve, hogy a fajok nem genetikailag egységesek. Ezek a vonalak evolúciósan jelentős egységekként (esu-k) jelennek meg. Az 1-es és a 3-as fajra a családközi génáramlás javallott. Ebben az esetben az evolúciós függetlenség átmeneti lehet, mivel ugyanazon fajhoz tartozó két vonal nem reprodukálható. Miközben különálló vonalként (fajként) maradnak, interspecifikus hibridizáció fordulhat elő a fajok közötti korlátozott géncserével (színváltozás).
Végrehajtása hatékony természetvédelmi stratégiák, amelyek célja, hogy megvédje mindkét faj genetikai sokszínűség előnyeit mind következetesebb nézet, hogy mi behatárolják faj, illetve más egységek szerte a lakosság-fajok kontinuum, hogy ezek az egységek értelmezni a különböző jogszabályok világszerte. A genomika új képességei mind lehetőségeket, mind kihívásokat nyújtanak e célok eléréséhez. A következőkben figyelembe vesszük: (én), hogy genom-skála elemzések össze lehet egyeztetni a konzervatív megközelítés, hogy a korlátozás, illetve elnevezése a fajok, míg a szintén azonosító fő összetevője a genetikai sokféleség belül a faj, (ii.), hogy a meglévő természetvédelmi jogszabályok az egész több kontinensen, hogy meghatározza, hogy a “fajok” közé széles infra-konkrét kategóriák közül, lehet használni, mivel a nagy felbontású által kínált genom képernyők, valamint (iii.), hogy a betekintést speciation folyamatok, valamint a szerkezetátalakítási variáció a genomika összehangolása, a jogszabályoknak, akkor a menedzsment. Ezután javasolunk néhány módot a védelmi egységek jogalkotási megközelítésekkel való összehangolására annak érdekében, hogy megvédjük a sokféleség teljes spektrumát a populációktól a fajokon keresztül.
Genomika, Speciation, valamint a Taxonómia
Mint sok területen a biológiai tudomány, rendszertan, a gyakorlat elnevezése taxonok, megértés a kapcsolatok, valamint milyen alkotnak, hogy jelentősen előrehaladott által genomika, statisztikai modellek. Akár teljes genomszekvenáláson keresztül (például Nater et al., 2017) vagy genom mintavétel (SNPs, target capture-szekvenálás; Jones and Good, 2016; Leache and Oaks, 2017), most már példátlan felbontása minták genetikai sokféleség (Funk et al., 2012). Párosulva egyre erősebb statisztikai modellek következtetett történetek genomikai illetve fajok divergencia, ezeket az adatokat nyújtó új betekintést az evolúciós folyamatok generálása faj genetikai variáció.
Ezen új eszközök alkalmazása két ellentétes betekintést generál. Egyrészt, sok tanulmány felfedi nagyon eltérő genetikai populációk belül megnevezett Fajok, úgynevezett rejtélyes Fajok (Bickford et al., 2007; Jorger and Schrödl, 2013; Struck et al., 2017). Ez semmi új, hiszen a térbeli populációgenetika és a molekuláris filogeográfia hosszú történetére épül, amely három évtizede produktív törekvés (Avise et al., 1987). A genomika mind a népességszerkezet, mind az adatok megnövekedett felbontása, amelyek elegendőek a népességtörténetek statisztikailag történő következtetéséhez és a divergencia alternatív modelljeinek teszteléséhez (Degnan and Rosenberg, 2009; Sukumaran and Knowles, 2017). Másrészt a genom-skála elemzések is sokatmondó, hogy (mint régóta ismerik a növényeket), genetikai csere (introgression) között állatfaj gyakoribb, mint azt korábban gondolták, mind során, illetve azt követően speciation, sőt, vezess új adaptív sugárzás (Mallet, 2007; Rieseberg and Willis, 2007; Arnold Kunte, 2017) 1. Ábra. Összerakva, ezek a betekintések tovább hangsúlyozzák, hogy a speciáció elhúzódik (Roux et al., 2016) és felfedi a phylogeography-filogenetics continuum (Edwards et al., 2016).
hogyan kapcsolódik mindez a taxonómiai és természetvédelmi célokhoz? Erős és igazolható aggodalom, hogy a genom képernyők által nyújtott fokozott felbontás a fajok burjánzó túlzott felosztásához vezethet (Isaac et al., 2004), potenciálisan korlátozva a gazdálkodási rugalmasságot, és a kis genetikailag eltérő populációkat beltenyésztésre és esetleges kihalásra (Frankham et al., 2012). Ezzel szemben, Gippoliti et al. (2018) azzal érvelnek, hogy a rendszertani tehetetlenség, amely korábban számos afrikai patás állatfaj felismerésének elmulasztását eredményezte, kitörési depressziót okozott a keverés után, valamint a földrajzilag korlátozott taxonok védelmének elmulasztása. Nézetük (és lásd még Morrison et al., 2009) az, hogy nincs bizonyíték arra, hogy a rendszertani infláció negatív hatással van a patás állatok védelmére (Gippoliti et al., 2018). A kapcsolódó aggodalom az, hogy az instabil taxonómia késlelteti mind a megőrzési gyakorlatot, mind a közvélemény észlelését (Garnett and Christidis, 2017). Ezeket az aggodalmakat részben a fajfogalmak és a szisztematikus biológia diagnosztikai módszerei körüli hosszú vita vezérli, különösen a filogenetikai Fajok fogalmának alkalmazása, amely szerint a fajok minimálisan diagnosztizálható egységek (Cracraft, 1983). A taxonómusok többsége azonban természeténél fogva konzervatív, csak a fajok elnevezése, ha a bizonyítékok több sorában egyetértés van (Dayrat, 2005; Padial et al., 2010; Schlick-Steiner et al., 2010: Yeates et al., 2011). Álláspontunk szerint a fajoknak olyan robusztus, egymástól függetlenül fejlődő vonalakat kell képviselniük, amelyek a közeli hozzátartozókkal való érintkezéskor nagyrészt érintetlenek maradnak-az evolúciós faj koncepciójának (Wiley, 1978) és az általánosított Vonalfaj-koncepciónak megfelelően (de Queiroz, 1998, 2007). A gyakorlatban ez közvetlen bizonyítékot igényel a reproduktív elszigeteltségről vagy ugyanazon ésszerű helyettesítőkről, miközben lehetővé teszi a korlátozott genetikai cserét (Singhal et al., 2018).
Genomikai adatok biztosít nagyobb hatalom, mint azelőtt, hogy azonosítsa a differenciálás, a divergencia belül, illetve fajok között, amely megköveteli, hogy nagyobb figyelmet, hogy ez miként hat körvonalazása a rendszertani egységek. Ahol a populációk erősen genetikailag és fenotípusosan eltérnek a már megnevezett fajokhoz képest, erős eset van arra, hogy ezeket külön fajként nevezzük el. Ha a populációk erősen eltérnek fenotípusosan, kevés genomikus szétválasztással, akkor ítélet születik arról, hogy ezek a fenotípusos különbségek (i) örökölhetők-e, és (ii) valószínűleg érdemi reprodukciós elszigeteltséget okoznak-e. A genomvizsgálatok az ilyen helyzeteket korlátozott génáramlás tesztelésével oldhatják meg. A nagyobb kihívást jelentő kérdés az, ahol morfológiailag hasonló, de genetikailag eltérő populációkat észlelnek. Az ilyen populációk könnyen diagnosztizálhatók különálló evolúciós vonalként statisztikai elhatárolási módszerek alkalmazásával a multilocus adatokra (Carstens et al., 2013; Rannala, 2015), de vajon ideiglenes populációk vagy tartós Fajok (Sukumaran and Knowles, 2017)? A nómenklatúra szabályain belül minden bizonnyal lehetséges, hogy a fajokat csak a DNS-szekvenciákból ismerjük fel, szekvencia – alapú diagnosztikát biztosítva (Tautz et al., 2003; szakács et al., 2010). Például Murphy et al. (2015) A DNS-szekvencia adatai alapján hat Ausztrál sivatagi tavaszi amphipod-fajt határoz meg és ír le, annak ellenére, hogy nincs egyértelmű morfológiai különbség. Ez azonban egy általános fejlesztési terület, ahol a rendszerezők a “rejtélyes Fajok” kifejezést nagyobb következetességgel és szigorúsággal kell alkalmazni (Struck et al., 2017).
azokban az esetekben, amikor a DNS-szekvencia-adatok potenciális kulcsfontosságú meghatározó tulajdonságot nyújtanak egy faj felismeréséhez, javasoljuk, hogy különösen fontos más bizonyítékok alkalmazása a túlcsordulás elkerülése érdekében (Oliver et al., 2015). Az egyik megközelítés a jelentős reprodukciós elszigeteltség tesztelése, ahol az ilyen populációk természetes érintkezési zónákat alkotnak. Például a fenotípusosan megkülönböztethetetlen Carlia gyíkok vonalpárjai közötti érintkezési zónák összehasonlító genomikai elemzésében a genetikai izoláció mutatói erősen korreláltak a divergencia idővel (Singhal and Moritz, 2013; Singhal and Bi, 2017) (2.ábra). Ezeknek a származáspároknak az eltérőbb változatosságát hivatalosan különálló fajként ismerik el, míg az érintkezési zónák közötti genomikusan kiterjedt genetikai csere bizonyítékaival szorosabban összefüggő (mégis statisztikailag diagnosztizálható) vonalak nem voltak (Singhal et al., 2018). Mivel az erős empirikus kapcsolat közötti eltérés, valamint a reproduktív izoláció ezek a fajok, ezek a szerzők további elválasztott két allopatric rejtélyes izolátumok (közeli rokon fajok), hogy volt még magasabb genomikai divergencia, mint azok, tagolt felett. A reprodukciós izoláció kiterjesztése más, mélyen eltérő, de morfológiailag hasonló filogeográfiai vonalakkal rendelkező kongener fajokra (Potter et al., 2016, 2018; Afonso Silva et al., 2017A) azt sugallja, hogy még több fajt kell leírni, különösen a szigeteken (2. ábra, Carlai amax és C. rufilatus), sőt, néhány ilyen fajt kiegészítő fenotípusos elemzés (Afonso Silva et al., 2017b). Más vonalak segítségével újabb eltérések, de még mindig statisztikailag tagolt, mint különálló vonalak segítségével a nagy készlet nukleáris gének, lehet képviseli, mint Evolúciós szempontból Jelentős Egységek (ESUs) belül faj.
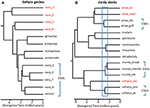
2.ábra. Az Ausztrál monszun trópusokból (AMT) származó gyíkok fajhatárainak filogenomikus felbontása. Minden esetben a starbeast2-ben megvalósított többfajtás coalescent segítségével 100 exonról becsülik meg a kapcsolatokat és a relatív eltérési időket, de a két példa közötti időskálák nem feltétlenül azonosak. A) A Gehyra Gekkos (Moritz et al., 2018). A G. nana-n belüli vonalvonalak filogenetikai sokfélesége, amint azt korábban felismerték, négy másik, már leírt és morfológiailag megkülönböztető fajt tartalmazott. A tartomány átfedéseinek és morfológiájának későbbi elemzése azt eredményezte, hogy a filogenetikailag leginkább eltérő vonalakat (vörös taxoncímkék) külön fajként (Doughty et al., 2018). A ma már elismert G. nana-ban maradt (morfológiailag elválaszthatatlan) vonalak esu-knak tekinthetők. B) A Fajok határainak folyamatos felbontása Carlia-ban az AMT-től. Egyes vonalakat már a genetikai Fajok elhatárolása és a diagnosztikai morfológiai különbségek (C. insularis és C. isostriacantha) post hoc azonosítása alapján fajként ismertek el; Afonso Silva et al., 2017b). Más komplexumok (Potter et al., 2016, 2018), rendszertani revíziók folyamatban vannak. A kék téglalap azt a divergenciaidő-tartományt jelöli, ahol a rokon fajok morfológiailag rejtélyes vonalai közötti érintkezési zónákban (Singhal and Bi, 2017) jelentős, a divergenciával korreláló reprodukciós izolációt figyeltek meg. Konzervatív, majd következő (Singhal et al., 2018), jelölt fajként három genetikai vonalat ismerünk el, amelyek eltérései meghaladják ezt az empirikus küszöböt. Ez genetikailag elkülönülő esu-kat hagyna Carlia amax, Carlia munda és Carlia rufilatus (kék zárójelben) között. A skála sávok becsléseket divergencia idő millió év.
szélsőséges esetekben a genomikai módszerek azt mutatják, hogy az egyetlen taxonómiai faj nemcsak genetikailag heterogén, hanem filogenetikai értelemben más elnevezett fajokat is tartalmaz. Az ilyen” paraphyly” akkor fordulhat elő, ha a közelmúltban egyetlen, genetikailag felosztott fajból (Patton and Smith, 1994) származik, de azt a hipotézist is generálja, hogy a genetikailag heterogén taxon több fajt képviselhet. Ismét több bizonyítékra van szükség. Ebben az esetben az észak-ausztráliai gehyra gecko gyíkok komplexének filogenomikai elemzése kilenc evolúciós vonalat határolt, köztük négy már elnevezett faj filogenetikailag beágyazva (Moritz et al., 2018; 2.ábra). A földrajzi átfedés későbbi bizonyítékai (a filogenetikai szétválasztás megtartása mellett) és a finom morfológiai különbségek alátámasztották a négy leginkább filogenetikailag eltérő vonal megnevezését (Doughty et al., 2018). Ezzel szemben a több, egymással szorosan összefüggő (még statisztikailag elkülöníthető) vonalak segítségével volt szomszédos ahelyett, hogy egymást átfedő disztribúciók, de nem voltak morfológiailag diagnosable, s ennek megfelelően megőrzött, mint egyetlen faj (G. nana értelemben stricto), bár több ESUs, a vizsgálat lezárásáig (2.Ábra). A minták későbbi genomvizsgálata ezen esu-k határain keresztül kimutatta, hogy legalább néhánynak jelentős genetikai cseréje van, az eredeti validálása, a fajok felismerésére alkalmazott konzervatív megközelítés.
egyéb esetekben növény genomikai vizsgálatok, még egyszerű koordinációs SNPs, felfedi stark, s váratlan eltérések között lakosság mintát, ami arra utal, hogy a jelenléte titokzatos taxonok, amelyek aztán lehet elemezni, diagnosztikai vizsgálatok, karakterek. Például, SNP-alapú bizonyítékok genomi divergencia populációk között, és ekotipikus különbségek eukaliptusz salubris kiderült két különböző molekuláris vonalak, hogy fenntartották a megkülönböztető területén földrajzi átfedés, az ökotípusos adaptáció tekinthető fontos tényező minimalizálásában gén áramlását a vonalszakaszok között. (Steane et al., 2015; 3. Egy másik esetben, az Ausztrál Muskátli, genomiális elemzés, kombinált morfológiai, illetve a reprodukciós vizsgálatok, oda vezetett, hogy jelentős változások taxonómia, mind a hasító -, olyan, mint a taxonok, különösen ott, ahol korábban a morfológiai tulajdonságok vagy megállapították, hogy bizonyíthatóan disszonáns a törzsfejlődés (Nicotra et al., 2016).
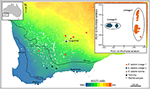
3. ábra. Genomikai elemzés segítségével SNP adatok azonosított rejtélyes vonalak a széles körben elterjedt eukaliptusz salubris. Az egyes leszármazások mintapopulációinak helyét a szárazsági index gradiensén mutatják Délnyugat-Nyugat-Ausztráliában. A 16,122 DArTseq markerből származó bináris genetikai távolságokon alapuló PCoA két különálló vonalat mutat. Adaptált Steane et al. (2015).
a fenti esetek szemléltetik az integratív taxonómiában (Padial et al., 2010; Yeates et al., 2011). Továbbá, ezek az esetek a genom redukciós szekvenálás valamilyen formáját használják, hogy kiváló minőségű adatokat szerezzenek az 1000 független génekből, ami sokkal nagyobb bizalmat ad az eredményekben,mint a csak plastid loci vagy néhány nukleáris gén felhasználásával. Példa arra, hogy az integratív taxonómia alkalmazása drámaian megváltoztathatja a sokféleség észlelését a Madagaszkári békák kombinált genetikai, akusztikus és morfológiai felméréséből származik, amely majdnem megduplázta az ismert fajok sokféleségét (Vieites et al., 2009). Ezek a szerzők a taxonómiai rendezés és kongruencia állapotát tükröző kifejezések sorozatát javasolták: Megerősített jelölt Fajok, genetikailag és fenotípusosan eltérő, de még nem nevezett taxonok esetében; meg nem erősített jelölt Fajok a genetikailag eltérő taxonok esetében, ahol a morfológiát és az akusztikát még nem vizsgálták; és mélyen specifikus vonalak az olyan entitások számára, amelyekről ismert, hogy genetikailag eltérnek, mégis fenotípusosan rejtettek. A genomikai elemzéseket még nem kell alkalmazni ezekre a taxonokra, de biztosan tájékoztatná a határozatokat a konspecifikus vonalokról vs.jelölt Fajok.
a genomikus megközelítések, még akkor is, ha konzervatív módon alkalmazzák, kétségtelenül a morfológiailag konzervatív taxonok további felosztásához vezetnek. De fordítva, vannak egyértelmű példák, ahol ugyanezek a megközelítések feltárják a túlzott osztott vagy helytelenül diagnosztizált fajokat (Nicotra et al., 2016). Természetesen már számos molekuláris tanulmány létezik a kisebb léptékű adatkészleteken alapuló növény-és állatfaj-komplexekről (pl., mikroszatellit, mtDNA és cpDNA), amelyek megállapították, hogy a fajokat tévesen diagnosztizálták, vagy ahol több fajt taxonómiailag körülhatároltak, de most egyetlen fajnak tekintik. Például Rabosky et al. (2014) használt mitokondriális és nukleáris gén filogenezis, hogy vizsgálja felül a scincid Ctenotus inornatus komplex, ami a nettó csökkenése fajok és átrendeződése Fajok határok; a morfológiai vonások korábban alkalmazott, míg kényelmes terepi biológusok, voltak nagyon labilis, és nagyrészt nem felel meg a fajok határait diagnosztizált genetikailag. A növények egy részét a legismertebb példák találhatók az orchideák, mint például az Európai orchidea nemzetség Ophrys, valamint az Ausztrál orchidea nemzetség Caladenia, ahol a molekuláris vizsgálatok azt mutatják, hogy jelentősen kevesebb szervezetek lenne ismerni, mint faj, mint azok alapján morfológiai tulajdonságok, valamint pseudocopulatory beporzás szindrómák (lásd Devey et al., 2008; Swarts et al., 2014).
Genom-skála adatok, ha konzervatív és ideális esetben a fenotípusok párhuzamos elemzésével értelmezik, továbbra is taxonómiai változásokhoz vezetnek, különösen a morfológiailag konzervatív taxában (Fišer et al., 2018). De ellentétben a egyesek által kifejezett aggodalmakkal (Garnett and Christidis, 2017), úgy gondoljuk, hogy ez az evolúciós szisztematika megfelelő törekvése, még akkor is, ha az eredmények ideiglenesen kényelmetlenek.
Genomics and Intraspecific Conservation Units
bár kevésbé vitatott, mint a taxonómiai elhatárolás és a fajszintű vita, de sok szempontból ugyanolyan összetett, volt a jelentős vita a meghatározása és elhatárolása természetvédelmi egységek fontos elemei a fajspecifikus sokféleség, hogy figyelembe kell venni a természetvédelmi intézkedések (Ryder, 1986; Waples, 1991; Dizon et al., 1992; Moritz, 1994; Vogler and de Salle, 1994; Crandall et al., 2000; Fraser and Bernatchez, 2001; Avise, 2004; Funk et al., 2012) (4. Tekintettel a fajok genomikai bizonyítékokból történő felismerésének óvatos megközelítésére, amelyet fent támogatunk (Lásd még Oliver et al., 2015; Singhal et al., 2018), elkerülhetetlen, hogy egyes rendszertani Fajok továbbra is hosszú, elszigetelt populációkat tartalmazzanak bennük. Olyan fogalmakat javasoltak, mint az evolúciósan jelentős egységek (esu-k) vagy a tervezhető egységek (du-k) a meglévő taxonómia kiegészítésére (Ryder, 1986; Waples, 1991; Moritz, 1994; Mee et al., 2015). A fajokhoz hasonlóan az értelmezések és a kritériumok is eltérőek, de a közös szándék az intraspecifikus sokféleség főbb elemeinek felismerése a természetvédelmi tevékenységek számára (Funk et al., 2012). Moritz (1994, 2002) támogatta egy egyszerű, de gyakorlatias molekuláris megközelítés azonosítása, egymástól függetlenül fejlődő szegmens belül a faj, a “történelmi” tengely a sokszínűség; míg mások több hangsúlyt az “adaptív” tengely közé funkcionális diverzitás (Ryder, 1986; Crandall et al., 2000; Fraser and Bernatchez, 2001; Waples et al., 2001; Mee et al., 2015) (4. Bármi legyen is ezeknek az alternatíváknak a relatív érdeme, egyértelmű, hogy a populációs genomika technikai és analitikai eszközei most már megalapozottan azonosíthatják az intraspecifikus sokféleség mindkét összetevőjét (Funk et al., 2012; Hoban et al., 2013; Whitlock, 2014; Yang and Rannala, 2014).
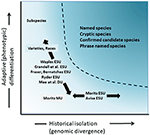
4.ábra. A genetikai sokféleség szétválasztása két összetevőre: adaptív variáció, amely a természetes szelekció révén keletkezik, és a fenotípusok és a történelmi elszigeteltség vagy a genomi divergencia által értékelt semleges divergencia elemzése. A görbe feletti sötétebb árnyékolt terület olyan körülményeket jelez, ahol a populációk valószínűleg különálló fajoknak tekinthetők a legtöbb fogalom alatt. A genetikai sokféleség két tengelyéhez viszonyítva (Moritz után, 2002) különböző természetvédelmi egységkoncepciókat és infra-specifikus taxonómiai kategóriákat mutatnak be.
Ez arra a kérdésre vezet, hogy miért nem csak az ESUs-t nevezik alfajnak? Az alfajokat hagyományosan mind a botanikusok, mind a zoológusok fenotípusosan elkülönülő, allopatric populációcsoportoknak tekintik, amelyek földrajzi határokon átterjedhetnek egymásra (Mayr, 1942; Grant, 1981). Mint ilyen, potenciálisan tükrözik az adaptív divergenciát anélkül, hogy szükségszerűen történelmileg elszigeteltek lennének. Az alfajok megnevezéseit széles körben használják a növényekben, földrajzi és ökológiai különbségekkel az elsődleges eszközök, amelyekkel megkülönböztetik őket (Hamilton and Reichard, 1992). Amennyiben alfaj nyilvánvaló helyi alkalmazkodás, vagy anélkül folyamatban lévő gén áramlását, azt javasoljuk, hogy phenotypically meghatározott alfaj továbbra is elismert, a természetvédelmi, valamint nem összetévesztette ESUs, legalább nem ESUs meghatározott képviseletére történelmileg elszigetelt populációk.
ugyanúgy, mint a lakosság genomika rendelkezik, sokkal nagyobb pontossággal azonosítása ESUs kihasználva mind a semleges, valamint az adaptív loci is biztosít azt jelenti, hogy kihasználva a nagyszámú semleges loci, hogy könnyebben behatárolják Igazgatási Egységek (MUs), mint a demográfiailag független egység jellemzi korlátozott gén flow (Funk et al., 2012). Míg a legtöbb itt a hangsúly az ESU és hasonló egységek, mint a DUs, hogy lehet elismerni, mint a fő intra-specifikus egységek megőrzése, és elfogadták a védelem egyes joghatóságok, MUs is azonosították képviselő demográfiai szempontból független populációk (lásd Palsbøll et al., 2007), hogy meg lehet őrizni a nagyobb ESU (Moritz, 1994; Funk et al., 2012). A gazdálkodási egységek fontosak lehetnek a fajpopulációk nyomon követésében és kezelésében, és “állományoknak” is nevezik őket, amelyek különösen fontosak a halászati gazdálkodás szempontjából (Avise, 2004; Laikre et al., 2005). A meghatározás MUs által Moritz (1994), mint a “lakosság jelentős eltérés az allél-gyakoriságok a nukleáris vagy mitokondriális loci, függetlenül attól, hogy a filogenetikai szól az allélek” általában széles körben elfogadott, bár, mint rámutatott Palsbøll et al. (2007), a demográfiai kapcsolat a legfontosabb tényező, nem pedig a történelmi génáramlás szintje.
a természetvédelmi egységeket az intraspecifikus sokféleség fontos elemeinek elhatárolása szintén jelentős hatással van a kis beltenyésztett populációk genetikai megmentésére a génáramlás helyreállítása vagy augmentáció révén, és egyre világosabbá válik, hogy a genetikai mentést szélesebb körben kell figyelembe venni, ha a népesség fokozott kihalását el kell kerülni (Love Stowell et al., 2017; Rals et al., 2018). Moritz (1999) javasolta, hogy a MUs keverése, de az ESUs nem tekinthető megfelelő stratégiának a genetikai mentéshez. Amint azt Frankham et al. (2012) a taxonómiailag elismert Fajok keverését általában nem tartják elfogadhatónak a vezetők, ebben az esetben a taxonómiai túltelítettség korlátozhatja a kis beltenyésztett populációk megmentésének lehetőségeit. Javasoljuk, hogy míg a szigorúbb megközelítés körülhatárolásáról fajon belüli egységek, mint például MUs segíthet enyhíteni ezt a problémát, az is egyértelmű, hogy genomikai becslési a migráció árak, valamint az adaptív eltérés van a potenciállal rendelkezik, hogy sokkal erőteljesebb útmutatást nem csak meghatározó fajok határait, valamint fajon belüli változatosságot, de a tervezés genetikai mentési stratégiák.
Filogenomika és természetvédelem
a filogenetikai módszerekkel kombinált genomika könnyen megoldhatja a taxonok közötti kapcsolatokat és evolúciós távolságot, valamint a fajok elhatárolását. Ezt legjobban a fajfa módszereivel lehet elérni, nem pedig a konkatenációval, mivel ez a génfa heterogenitása, valamint az ősi polimorfizmus (Edwards et al., 2016; Ogilvie et al., 2016). Míg a loci nagy száma gyakran nem nélkülözhetetlen, beleértve a több loci-t is, gyakran javítja mind a kapcsolatok, mind az ág hosszának felbontását (pl., Blom et al., 2016). A megőrzés fontossága kettős. Először is, a filogenetikai értékelések lehetővé teszik a fajok vagy az esu-k megkülönböztethetőségének figyelembevételét, nem csak arról, hogy leírják-e őket. Így az EDGE program (Isaac et al., 2007) támogatja a fajok megőrzésének súlyozását annak alapján, hogy a szokásos IUCN kritériumok mellett mennyi egyedi evolúciós történetet képviselnek. Második, ág hossza maguk képviselik a biodiverzitás értékek, független a taxonómia (Hit, 1992), amely bebizonyította, érték előtérbe területek természetvédelmi (Laikusok et al., 2015; Rosauer et al., 2016, 2018). Ebben az értelemben a filogenetikai metrikák elfogadása olyan intézkedések alkalmazásával, mint a filogenetikai sokféleség és a filogenetikai endemizmus, lehetővé teszi a magas evolúciós sokféleség olyan területeinek kiemelését, amelyek védelemre irányulhatnak, és kiemelt prioritást élveznek a biológiai sokféleség jobb megőrzésének bármely tervezési folyamatában. Ez a megközelítés fokozhatja a genetikai sokféleség és az evolúciós folyamatok védelmét az organizmusok különböző csoportjai számára számos földrajzi skálán (Laity et al., 2015) A területekkel kapcsolatos döntések meghozatala annak érdekében, hogy a genomikai bizonyítékoknak a fajok vagy a fajon belüli egységek elismerésére történő lefordításának bizonytalanságaira támaszkodjanak.
A valós világ-jogalkotási fogalommeghatározások
a biológiai sokféleség megőrzésére vonatkozó politikai és jogszabályi keretek a nemzetközi egyezményektől és stratégiáktól, a nemzeti stratégiákon és jogszabályokon, valamint az országokon belüli állami vagy regionális jogszabályokon és stratégiákon keresztül terjednek. A nagy nemzetközi eszközök a biológiai védelmi közé tartozik a Biológiai Sokféleség Egyezmény végrehajtása révén az Egyezmény a Felek (COP), a Világörökség Egyezmény, a Globális Biodiverzitás Stratégia, a Montreali Folyamat az Erdő Védelméről, a Ramsari Egyezmény a Vizes élőhelyek, az Egyezmény a Vándorló Fajok, az Egyezmény a Nemzetközi Kereskedelem a Veszélyeztetett Fajok (CITES), az International Union for Conservation of Nature (IUCN) vörös lista a veszélyeztetett fajok, valamint az Egyesült Nemzetek szervezete éghajlat-változási keretegyezménye. Mindezek az oklevelek a fajok megőrzésének és a genetikai sokféleség megőrzésének fő célját testesítik meg, mint a biológiai sokféleség megőrzésének alapvető célját, a fajok pedig a védelem széles körben meghatározott egysége. A fajok jogi és nemzetközi irányelvek szerinti meghatározása azonban rendkívül változó. Ezek magukban foglalják a fajok taxonómiai elismerését és az intraspecifikus taxonokat, de tartalmazhatnak olyan genetikai/evolúciós fogalmakat is, amelyek gyakran egy “faj” jogi meghatározása alá tartoznak (1.táblázat).

1. táblázat. A világ jogszabályai és irányelvei, amelyek “fajokat” határoznak meg a jegyzékbe vétel és a védelem tekintetében.
tekintettel a védhető infra-specifikus szintek széles körére (1.táblázat), az országok jogszabályainak és kapcsolódó jegyzékbe vételének megkönnyítenie kell a genetikai sokféleség megőrzését. Például egy “faj” meghatározása, amikor a világ minden tájáról figyelembe vesszük a jogszabályokat (1. táblázat), a következőket foglalhatja magában: fajokat, alfajokat, fajtákat, földrajzilag és/vagy genetikailag elkülönülő populációkat, míg a Dél-afrikai jogszabályok értelmében a fajtákat, földrajzi fajokat, törzseket és hibrideket is magában foglalhatja. Így a legtöbb jogszabály szerint globálisan egy ” faj ” olyan entitásokat foglal magában, amelyek különböző infra-specifikus szinteket tükröznek, amelyek mind egy szélesebb fajeloszlás részét képezik, és különböző mértékben tükrözik a populáció-faj kontinuumot. Ez párhuzamot von azzal az elkerülhetetlen nehézséggel, amellyel a taxonómusok, az evolúciós biológusok és a természetvédelmi genetikusok szembesülnek abban, hogy következetesebb és egységesebb módszert találjanak a kontinuumon belüli egységek meghatározására.
egyesek azzal érvelnek, hogy a fajok politikai következményeivel és az infra-specifikus bizonytalansággal kapcsolatos aggodalmak nem megfelelőek (Hey et al., 2003), de még mindig az a nézet, hogy míg a szabályozás biztosítja, hogy egy mechanizmus védi a genetikai sokszínűség, a sokféle szint, a bizonyítás terhe is biológusok további világosság, állaga, hogy ezeket a szinteket kell tagolt a faj át, hogy az egyes populációk. Jelenleg a vezetők, valamint a jogalkotók maradt egy barátságtalan tömb faj fogalmak, illetve infra-speciális egységek által nyújtott tudósok meghatározásakor egység át a lakosság-fajok folytonosság. Például, minden jogrendszerben fontolja meg fajok által elismert taxonomists, mint kulcs egység megőrzése (1. Táblázat), de még mindig széles körű vita között taxonomists körül faj koncepciók, valamint hogy a fajok hogyan diagnosztizálják a gyakorlatban változik belül, illetve az egész csoport. Ez ahhoz vezetett, hogy a közelmúltban felhívták a taxonómia szabályozottabb megközelítését (Garnett and Christidis, 2017), amelyre mások ellenezték, hogy a taxonómia szabályozása nem a válasz (Thomson et al., 2018). Bár egyetértünk néhány pontot tett Thomson et al. (2018) Azt javasoljuk, hogy meglehetősen naivitás azt javasolni, hogy a jogszabályoknak meg kell változniuk ahhoz, hogy kifejezetten hivatkozzanak a név által feltételezett konkrét taxonkoncepcióra. Ezek, valamint a korábbi ilyen viták hangsúlyozni szeretném, hogy az evolúciós biológusok, valamint taxonomists, hogy átvegye a vezetést az egyértelműség biztosítása (átláthatóság), hogy a fordítás genomikai illetve egyéb bizonyítékot, hogy lehatárolása faj vagy fajon belüli egységek, valamint, hogy ezek a fajok vonatkoznak vonatkozó jogszabályok védelme, valamint.
fordítás menedzsment
amint azt fentebb tárgyaltuk, minden jogszabály és irányelv a fajszintet a megőrzés alapvető egységeként ismeri el, de biztosítja a különböző infra-specifikus egységek elismerését is. Sokan a faj hivatalos elnevezésére és taxonómiai leírására is hallgatólagos követelményt fogalmaznak meg, ha természetvédelmi prioritásként és formális védelemként tekintenek rájuk. Például az IUCN Red Listing Guidelines szerint a meg nem nevezett Fajok felsorolása nem ajánlott, és követelmény, hogy az új fajleírást négy éven belül közzé kell tenni, miután a faj szerepel az IUCN vörös listáján, vagy el kell távolítani. Ez kiemeli annak fontosságát, hogy taxonómiai elnevezése, leírása szervezetek, mind a fajok, illetve infra-specifikus szintje, a korlátozott rendszertani információt a sok növényi, mind az állati csoportok (Mace, 2004; Oliver Lee, 2010; Wilson, 2017), valamint az, hogy gyorsítsa fel rendszertani erőfeszítést, különösen a biológiai sokféleség gazdag területeken (Riedel et al., 2013; Wege et al., 2015; Wilson, 2017).
Taxonomically nevű fajt, illetve fajon belüli rendszertani kategóriák, mint a faj, illetve fajta, általában biztosítja az alapvető egység a hivatalos lista, a természetvédelmi prioritások, valamint a védelem, még néhány jogrendszer más kategóriák fajon belüli védelmi egységek hivatalosan is elismert. Ezeket általában kombinált genomikai és fenotípusos adatok alapján azonosítják. Például az USA veszélyeztetett fajokról szóló törvény elismeri a különféle populációs szegmenseket (DPSs), amelyek széles körben az esu-kon alapulnak (Waples, 1991), míg a veszélyeztetett Kanadai fajokról szóló törvény elismeri A DUs-t. A “különálló populációk” operatív értelmezése a veszélyeztetett Kanadai fajokról szóló törvény szerint A DUs célja azon populációk diszkrét halmazainak azonosítása, amelyek a fajok evolúciós örökségének fontos alkotóelemeit képviselik, és amelyek valószínűleg nem helyettesíthetők természetes diszperzióval (Mee et al., 2015).
mind a DUs, mind a DPSs fogalmilag átfedésben van az esu-kkal (Waples et al., 2013), és a legszélesebb körben alkalmazták az észak-amerikai halakra, köztük az USA északnyugati részén található Lazacokra (Waples, 1995) és a kanadai coregonidákra (Mee et al., 2015) érdekes módon az USA-ban a DPSs csak a gerincesekre vonatkozik, míg Kanadában A Dus alkalmazható bármely olyan szervezetre, amely a törvény alapján felsorolható (lásd Waples et al., 2013), A DUs jelenleg szerepel ízeltlábúak, puhatestűek és növények. Ausztráliában, akárcsak Kanadában, az 1999.évi Környezetvédelmi és biodiverzitás-védelmi törvény (EPBC Act) alapján elismert bármely szervezet esetében fel lehet sorolni a különálló populációkat, bár ezek nem kifejezetten elnevezett kategóriák, mint például a DUSs és a DPSs. A mai napig azonban csak gerincesek számára ismerték fel őket. Az USA-BAN 127 gerinces DPSs van felsorolva a megőrzés céljából (https://ecos.fws.gov/ecp/); Kanadában 70 Du-t soroltak fel az összes szervezetre (http://www.registrelep-sararegistry.gc.ca/species/schedules_e.cfm?id=1); Ausztráliában pedig az emlősöket, halakat és madarakat lefedő 7 gerinces faj populációit az EPBC-törvény alapján különleges természetvédelmi státusszal ismerték el (http://www.environment.gov.au/cgi-bin/sprat/public/publicthreatenedlist.pl). Míg az Egyesült Államokban és Kanadában e populációk értékelése általában genetikai információt használ (Waples et al., 2013), Ausztráliában általában földrajzi és joghatósági azonosításon alapulnak, bár egyes esetekben az emlősök genetikai differenciálódásának elemzésével (Firestone et al., 1999; Armstrong, 2006) és fish (Colgan and Paxton, 1997; Ahonen et al., 2009).
A lista az infra-specifikus védelmi egységek növények gerinctelenek Kanadában (DUs), de nem az USA-ban, vagy Ausztrália rávilágít egy másik kérdés, nem kifejezetten kapcsolódik a lehatárolása védelmi egységek de szélesebb értelemben jelentős ellentmondások a biológiai sokféleség megőrzésére rangsorolása; ikonikus gerincesek, mint a madarak, emlősök, halak van a hangsúly sok jogrendszerben, míg más szervezetek, vagy nem. Az egyes szervezetek populációinak ez a preferenciális felsorolása számos olyan kérdést tükröz, amelyek kevésbé kapcsolódnak a természetvédelmi egységek határolásának tudományához, és inkább a populációk nyilvános tudatosság, politikai célszerűség és a felsorolt szervezet karizmatikus jellege alapján történő felsorolásához (lásd Waples et al., 2013).
Míg Ausztráliában egyes populációk jól ismert emlősök, madarak, halak védett alatt a EPBC Törvény, ott már jelentős mértékű információk alapján a molekuláris vizsgálatok hüllők, gerinctelenek, a növények pedig, ahol a lakosság is lehet elismert, mint ESUs vagy MUs, de sok ezek közül a természetvédelmi jelentősége, mivel a szűk földrajzi tartományban, a kis populáció mérete, aránya csökken, illetve érzékenység fenyegető folyamatokat. Például, feltűnő példa erősen struktúrált populációk mélyen eltérő vonalak segítségével belül faj, ami könnyen illeszkedjen a követelmény egy különleges megtalálható egy sor alacsony terjedési gerincesek a trópusokon (2. Ábra), de másutt is. Hasonlóan az Ausztrál gerinctelenek, mint amphipods, valamint százlábúak, jelentős genetikai szerkezetét, phylogeographic minták támogatja a lehatárolása diszkrét lakosság klaszterek, mint ESUs, amelyek földrajzilag különálló, keskeny földrajzi tartományok, kizárólag egyedi élőhelyek, valamint képviseli, csak néhány populációk (Finston et al., 2007; Murphy et al., 2013; Nistelberger et al., 2014a). Gyakran előfordul, hogy ezek a rövid hatótávolságú endemikusak a megőrzési aggodalomra okot adó taxonok (Harvey et al., 2011), valamint az endemizmus hotspotok (pl., 2017), amelyek maguk is kiemelt prioritások a több taxon megőrzési figyelem szempontjából (Rosauer et al., 2018), Mint például a Kimberley régió, valamint a Stirling tartomány délnyugat-Nyugat-Ausztráliában.
Az Ausztrál növények is vannak jelentős számú népesség genetikai, mind phylogeographic tanulmányok azonosítása különböző lakossági csoportok tartományon belül a faj a különböző nemzetségek tükröző disjunct, illetve történelmileg elszigetelt populáció rendszerek, földtani, edaphic bonyolult, de megszálló kontrasztos élőhelyek szempontjából növényzet, valamint az éghajlatváltozás (lásd a Broadhurst et al., 2017). A genetikailag különálló populációk különösen nyilvánvalóak a délnyugat-Ausztrália fajaiban (Coates, 2000; Byrne et al., 2014) számos tanulmányban kiemelve a jelentős genetikai struktúrát és a növénynemzetségek és a családok egy sor nagyon eltérő vonalát, többek között: Akác (Mimosaceae; Coates, 1988), Atriplex (Amaranthaceae; Shepherd et al., 2015), Banksia (Proteaceae; Coates et al., 2015), Lambertia (Proteaceae; Byrne et al., 2002), Hakea (Proteaceae; Sampson et al., 2014), Eremophila (Scrophulariaceae; Llorens et al., 2015), Pultanaea (Fabaceae; Millar and Byrne, 2013) és Calothamnus (Myrtaceae; Nistelberger et al., 2014b). Sok esetben ezeket a populációkat az állami kormányzati szervek a természetvédelmi tervezés során elismerik, és figyelembe veszik a környezeti hatásvizsgálatokban (lásd Shepherd et al., 2015) annak ellenére, hogy nem szerepelnek a jogszabályok szerinti védelemben. Elismerjük azonban, hogy ezeknek az egységeknek a formális elismerése hosszú távon javítaná azok megőrzését.
Ezek az Ausztrál példák rámutatnak arra, hogy egyre nagyobb szükség van a genetikai sokféleség ezen rétegének felismerésére a fajszint alatt. A kihívás annak biztosítása, hogy a természetvédelmi szempontból aggályos erősen strukturált populációk és mélyen eltérő vonalvonalak védve legyenek a természetvédelmi figyelem iránti versengő igényekkel szemben. Az ikonikus fajokon túl ez a kihívás világszerte is nyilvánvaló, Kanada kivételével. Hiba, hogy megvédje világosan körülhatárolt védelmi egységek valószínűleg növeli a rejtélyes kihalás veszteség jelentős genetikai sokszínűség, amely sok esetben járhat vonalak segítségével, illetve történelmileg elszigetelt populációk, amelyek kitartott belül a faj, több millió év (Moritz, 2002).
következtetések és ajánlások
a veszélyeztetett fajokra vonatkozó jogszabályok vagy nemzetközi szervezetek, például az IUCN szerint a fajok és a különböző infra-specifikus kategóriák felsorolása elkötelezettséget mutat a fajpopuláció-kontinuum genetikai sokféleségének védelme iránt. A biológiai sokféleség megőrzésének jelenlegi megközelítései azonban nagyrészt e kontinuum fajvégére összpontosítanak, kevesebb figyelmet fordítva a genetikai sokféleségre és az infra-specifikus változásokra. Ez a fajon belüli szintű genetikai változásokra való korlátozottabb összpontosítás azt sugallja, hogy míg a természetvédelmi vezetők egyre inkább felismerik a genetikai információ kifejezett megfontolásának értékét a gazdálkodási tevékenységekben és a prioritások meghatározásában, gyakran hiányoznak a biológiai sokféleséggel foglalkozó tudósok (Waples et al., 2008; Laikre et al., 2010; szakács és Sgrò, 2017; Taylor et al., 2017). Ez a tudományos és természetvédelmi gyakorlat közötti különbség akadályozza a genetikai sokféleség megőrzésének és kezelésének hatékonyabb globális megközelítését.
azt javasoljuk, hogy a biológiai sokféleség tudósainak a megőrzési egységekkel szembeni egyértelműségének hiánya nagyrészt két szinten fennálló következetlenségeken alapul. Az első a különböző fajfogalmakra és a különböző diagnosztikai módszerekre vonatkozik, amelyek a fajok körülhatárolására szolgálnak, ami következetlenségeket okoz a rendszertani csoportok között. A második a taxonómusok, az evolúciós biológusok és a természetvédelmi biológusok következetességének és egyetértésének hiányával kapcsolatos, hogy hogyan kell kezelni az infra-specifikus taxonokat és más szervezeteket, például az ESUs-t, A DUs-t és a MUs-t.
Amíg úgy gondoljuk, hogy a jogszabályok gyakran már kellően rugalmas, tekintve, hogy taxonok (vagy a “faj”) határozza meg, azzal érvelünk, hogy szükség van arra, hogy rendszeresen foglalkozzon a lakosság faj folytonosság, valamint több határozza meg egyértelműen egység megőrzése, nagyobb elismerés, tekintve, hogy olyan fogalmakat, mint a KOMMANDÓSOK, illetve, hogy ezek kapcsolódnak a vonatkozó jogszabályokat. Hangsúlyozzuk, hogy ez a kérdés kétségtelenül kritikusabbá válik, tekintettel arra, hogy a genomika képes a differenciálódást fokozatosan finomabb skálákon felismerni, és egyre inkább felhasználja a fajokat és az infra-specifikus entitásokat.
az előző elemzés, ajánljuk a következő ajánlásokat:
(i) Genomikai adatait kell értelmezni, konzervatív meghatározásakor faj, segítségével egy integratív rendszertani megközelítés, amely jellemzően magában foglalja a bizonyíték túl a genetikai adatok egyedül.
(ii
(iii.) fontolni az IUCN, a Természetvédelmi Genetika Specialista Csoport, valamint a regionális alcsoportok, hogy a fejlődő iránymutatások felismerve, valamint a címke fajon belüli egységek összehangolása, a legjobb tudományos gyakorlat.
(iv) A árnyaltabb megtekintése kialakuló evolúciós genomika a fajok, valamint a folytonosság a populációk, fajok kell közölni hatékonyabban a tudósok, hogy természetvédelmi vezetők, valamint a politikai döntéshozók.
tágabb értelemben a közelmúltban számos véleményben tett ajánlásokat visszhang (Frankham, 2010; Santamaría and Méndez, 2012; Hoban et al., 2013; Moritz and Potter, 2013; Laity et al., 2015; Pierson et al., 2016; szakács és Sgrò, 2017; Taylor et al., 2017), hogy hangsúlyozni szeretném, hogy a tudósok, hogy a kommunikáció javítása, valamint a tudatosság a politikai döntéshozók, a kormányzati szervek, a nem-kormányzati szervezetek, valamint a szakemberek, hogy az eredmények a molekuláris rendszertan, molekuláris taxonómia, phylogeography, valamint a lakosság genetikai kutatás adhat kritikus információ a genetikai sokféleség megőrzése, kezelése evolúciós folyamatok.
szerzői hozzájárulások
DC, MB, CM mind hozzájárultak a kézirat elkészítéséhez és felülvizsgálatához.
összeférhetetlenségi nyilatkozat
a szerzők kijelentik, hogy a kutatást olyan kereskedelmi vagy pénzügyi kapcsolatok hiányában végezték, amelyek potenciális összeférhetetlenségnek tekinthetők.
elismerések
ezt a projektet az Ausztrál Kutatási Tanács támogatta (ARC Linkage Grant No. LP120200063), az Ausztrál Nemzeti Egyetem és a nyugat-ausztrál biodiverzitás, természetvédelem és látnivalók Tanszék.
Afonso Silva, A. C., Bragg, J. G., Potter, S., Fernandes, C., Coelho, M. M., and Moritz, C. (2017A). Trópusi specialista versus climate generalist: diverzifikáció és demográfiai története testvér Fajok Carlia skinks Északnyugat-Ausztráliában. Mol. Ecol. 26:4045–4058. doi: 10.1111 / mec.14185
CrossRef Full Text / Google Scholar
Afonso Silva, A. C., Santos, N., Ogilvie, H. A., and Moritz, C. (2017b). Két új, északnyugat-Ausztrál Szivárványbőr érvényesítése és leírása több fajból álló coalescent módszerekkel és morfológiával. PeerJ 5: e3724. doi: 10.7717 / peerj.3724
P > PubMed Abstract / CrossRef teljes szöveg | Google Scholar
Ahonen, H., Harcourt, R. G., and Stow, A. J. (2009). A nukleáris és mitokondriális DNS a megszemélyesített szürke cápapopulációk (Carcharias taurus) izolációját tárja fel. Mol. Ecol. 18, 4409–4421. doi: 10.1111 / j. 1365-294x.2009.04377.x
PubMed Abstract / CrossRef teljes szöveg | Google Scholar
Armstrong, K. N. (2006). Phylogeographic structure in Rhinonicteris aurantia (Chiroptera: Hipposideridae): implications for conservation. Acta Chiropterologica 8, 63-81. doi: 10.3161/1733-5329(2006)82.0.CO
CrossRef Full Text / Google Scholar
Arnold, M. L., és Kunte, K. (2017). Adaptív genetikai csere: a keveredés és az evolúciós innováció kusza története. Trendek Ecol. Evol. 32, 601–611. doi: 10.1016 / j.fa.2017.05.007
P> PubMed Abstract | CrossRef teljes szöveg/Google Scholar
Avise, J. C. (2004). Molekuláris markerek, természettudományok és evolúció. A 2. Sunderland, MA; Oxford: Oxford University Press.
Google Scholar
Avise, J. C., Arnold, J., Ball, R. M., Bermingham, E., Lamb, T., Neigel, J. E., et al. (1987). Intraspecifikus filogeográfia: a mitokondriális DNS híd a populációs genetika és a szisztematika között. Ann. Rev. Ecol. Sys. 18, 489–522. doi: 10.1146 / annurev.es.18. 110187.002421
CrossRef teljes szöveg | Google Scholar
Bickford, D., Lohman, D. J., Sodhi, N. S, Ng, P. K., Meier, R., Winker, K., et al. (2007). Rejtélyes fajok, mint a sokféleség és a természetvédelem ablaka. Trendek Ecol. Evol. 22, 148–155. doi: 10.1016 / j.fa.2006.11.004
P>PubMed Abstract | CrossRef Full Text | Google Scholar
Blom, M. P. K., Bragg, J. G., Potter, S., and Moritz, C. (2016). Számviteli bizonytalanság gén fa becslés: összefoglaló-coalescent Fajok fa következtetés egy kihívást jelentő sugárzás Ausztrál gyíkok. Syst. Biol. 66, 352–366. doi: 10.1093 / sysbio / syw089
PubMed Abstract / CrossRef teljes szöveg / Google Scholar
Broadhurst, L., Breed, M., Lowe, A., Bragg, J., Catullo, R., Coates, D. J., et al. (2017). Az Ausztrál flóra genetikai sokfélesége és szerkezete. Búvárok. Distrib. 23, 41–52. doi: 10.1111 / ddi.12505
CrossRef Full Text / Google Scholar
Byrne, M., Coates, D. J., Forest, F., Hopper, S. D., Krauss, S. L., Sniderman, J. M. K., et al. (2014). “Változatos növény-és genetikai kapcsolatok”, a növényvilágban a Délnyugat-ausztráliai Homokmezőkön, a biológiai sokféleség globális Hotspotján, szerkesztők H. Lambers (Crawley, WA: University of Western Australia Publishing), 81-99.
P > PubMed Abstract
Byrne, M., Macdonald, B., and Coates, D. J. (2002). Phylogeographic patterns and high cpdns diversity within the Acacia acuminata (Leguminosae:Mimosoideae). komplexum Nyugat-Ausztráliában. J. Evol. Biol. 15, 576–587. doi: 10.1046 / j. 1420-9101.2002.00429.x
CrossRef Full Text / Google Scholar
Carstens, B. C., Pelletier, T. A., Reid, N. M., and Satler, J. D. (2013). Hogyan lehet kudarcot vallani a fajhatárolásnál. Mol. Ecol. 22, 4369–4383. doi: 10.1111 / mec.12413
P > PubMed Abstract / CrossRef teljes szöveg | Google Scholar
Coates, D. J. (1988). Genetikai sokféleség és populációs génstruktúra a ritka Chittering grass wattle-ben, Acacia anomala Court-ban. Aust. J. Bot. 36:273–286. doi: 10.1071/BT9880273
CrossRef teljes szöveg | Google Scholar
Coates, D. J. (2000). A gazdag és töredezett növényvilágban a természetvédelmi egységek meghatározása: a délnyugat-ausztráliai növények genetikai erőforrásainak és evolúciós folyamatainak kezelésére gyakorolt hatások. Aust. J. Bot. 48, 329–339. doi: 10.1071/BT99018
CrossRef teljes szöveg | Google Scholar
Coates, D. J., McArthur, S. L., and Byrne, M. (2015). A ritka endemikus Banksia brownii (Proteaceae) populációjának kórokozó által vezérelt kipusztulását követő jelentős genetikai sokféleség-veszteség. Biol. Conserv. 192, 353–360. doi: 10.1016 / j. biocon.2015.10.013
CrossRef Full Text / Google Scholar
Colgan, D. J., and Paxton, J. R. (1997). A rexea solandri (Scombroidea: Gempylidae) nevű közönséges gyöngyhal nyugati állományának biokémiai genetikája és felismerése., Ausztráliában. Mar. Freshw. Res. 48, 103-118. doi: 10.1071/MF96048
CrossRef teljes szöveg/Google Scholar
Egyezmény a biológiai sokféleségről (2007). Elérhető online: https://www.cbd.int/doc/meetings/cop-bureau/cop-bur-2007/cop-bur-2007-10-14-en.pdf
szakács, C. N., and Sgrò, C. M. (2017). A tudomány és a politika összehangolása az evolúciósan felvilágosult megőrzés elérése érdekében. Conserv. Biol. 31, 501–512. születési idő: 10.1111 / cobi.12863
P > PubMed Abstract / CrossRef teljes szöveg | Google Scholar
szakács, L. G., Edwards, R. D., Crisp, M. D., and Hardy, N. B. (2010). Szükség van morfológiára mindig az új fajleírásokhoz? Fordított. Sys. 24, 322–332. doi: 10.1071/IS10011
CrossRef teljes szöveg/Google Scholar
Cracraft, J. (1983). Faj fogalmak és speciációs elemzés. Curr. Ornithol. 1, 159–187. doi: 10.1007/978-1-4615-6781-3_6
CrossRef Full Text / Google Scholar
Crandall, K. A., Bininda-Emonds, O. R., Mace, G. M., and Wayne, R. K. (2000). Figyelembe véve az evolúciós folyamatokat a természetvédelmi biológiában. Trendek Ecol. Evol. 15, 290–295. doi: 10.1016 / S0169-5347 (00)01876-0
P> PubMed Abstract | CrossRef teljes szöveg | Google Scholar
Cutter, A. D., and Payseur, B. A. (2013). A szelekció genomikus aláírásai a kapcsolódó helyszíneken: a fajok közötti különbség egységesítése. Nat. Genet Tiszteletes. 14, 262–274. doi: 10.1038/Nrg3425
P> PubMed Abstract | CrossRef teljes szöveg/Google Scholar
Dayrat, B. (2005). Az integratív taxonómia felé. Biol. J. Linn. Soc. 85, 407–415. doi: 10.1111 / j. 1095-8312.2005.00503.x5
CrossRef Full Text / Google Scholar
de Queiroz, K. (1998). “The general lineage concept of species, species criteria, and the process of speciation,” in Forms: Species and Speciation, D. J. Howard, r S. H. Berlocher (New York, NY: Oxford University Press), 57-75.
Google Scholar
de Queiroz, K. (2007). Fajfogalmak és fajhatárolás. Syst. Biol. 56, 879–886. doi: 10.1080 / 10635150701701083
PubMed Abstract | CrossRef teljes szöveg | Google Scholar
Degnan, J. H., and Rosenberg, N. A. (2009). Gene tree discordance, filogenetikai következtetés és a multifajok coalescent. Trendek Ecol. Evol. 24, 332–340. doi: 10.1016 / j.fa.2009.01.009
P>PubMed Abstract | CrossRef Full Text | Google Scholar
Devey, D. S., Bateman, R. M., Fay, M. F., and Hawkins, J. A. (2008). Barátok vagy rokonok? Filogenetika és fajhatárolás a vitatott Európai orchidea nemzetség Ophrys. Ann. Bot. 101, 385–402. doi: 10.1093/AOB/mcm299
P>PubMed Abstract | CrossRef teljes szöveg | Google Scholar
Dizon, A. E., Lockyer, C, Perrin, W. F., Demaster, D. P., and Sisson, J. (1992). Az állománykoncepció újragondolása-filogeográfiai megközelítés. Conserv. Biol. 6, 24–36. doi: 10.1046 / j. 1523-1739. 1992. 610024.x
CrossRef Full Text / Google Scholar
Doughty, P., Bourke, G., Tedeschi, L., Pratt, R., Oliver, P., Palmer, R., et al. (2018). A faj a Gehyra nana (Squamata: Gekkonidae) alcsaládjába tartozik. komplex: rejtélyes és eltérő morfológiai evolúció az Ausztrál monszun trópusokon, négy új faj leírásával. Zootaxa, 4403:201-244. doi: 10.11646 / zootaxa.4403.2.1
P> PubMed Abstract | CrossRef teljes szöveg/Google Scholar
Dynesius, M., and Jansson, R. (2014). A fajon belüli vonalak perzisztenciája: a speciációs Arány elhanyagolt ellenőrzése. Evolúció 68, 923-934. doi: 10.1111 / evo.12316
P > PubMed Abstract / CrossRef teljes szöveg | Google Scholar
Edwards, S. V., Fazekas, S., Schmitt, C. J., Bragg, J. G., and Moritz, C. (2016). Retikuláció, divergencia, és a filogeográfia-filogenetika continuumc. Proc. NAT. Acad. Sci. U. S. A. 113, 8025-8032. doi: 10.1073 / pnas.1601066113
CrossRef teljes szöveg | Google Scholar
hit, D. P. (1992). Természetvédelmi értékelés és filogenetikai sokféleség. Biol. Hátrányok. 61, 1–10. doi: 10.1016/0006-3207(92)91201-3
CrossRef Full Text / Google Scholar
Finston, T. L., Johnson, M. S., Humphreys, W. F., Eberhard, S. M., and Halse, S. A. (2007). Rejtélyes speciáció két elterjedt föld alatti amphipod nemzetségek tükrözi történelmi vízelvezető minták egy ősi táj. Mol. Ecol. 16, 355–365. doi: 10.1111 / j. 1365-294x.2006.03123.x
PubMed Abstract / CrossRef Full Text / Google Scholar
Firestone, K. B., Elphinstone, M. S., Sherwin, W. B., and Houlden, B. A. (1999). Phylogeographical population structure of tiger quolls Dasyurus maculatus (Dasyuridae: Marsupalia), egy veszélyeztetett húsevő marsupial. Mol. Ecol. 8, 1613–1625. doi: 10.1046 / j. 1365-294x.1999.00745.x
CrossRef Full Text / Google Scholar
Fišer, C., Robinson, C. T., and Malard, F. (2018). Rejtélyes fajok, mint egy ablak a paradigmaváltás a faj fogalom. Mol. Ecol. 27, 613–635. doi: 10.1111 / mec.14486
P > PubMed Abstract / CrossRef teljes szöveg | Google Scholar
Frankham, R. (2010). A biológiai megőrzés genetikai megközelítésének kihívásai és lehetőségei. Biol. Conserv. 143,1919–1927. doi: 10.1016 / j. biocon.2010.05.011
CrossRef teljes szöveg | Google Scholar
Frankham, R., Ballou, J. D., Dudash, M. R., Eldridge, M. D., Fenster, C. B., Lacy, R. C., et al. (2012). A különböző fajok fogalmainak következményei a biológiai sokféleség megőrzésére. Biol. Conserv. 153, 25–31. doi: 10.1016 / j. biocon.2012.04.034
CrossRef teljes szöveg/Google Scholar
Fraser, D. J., and Bernatchez, L. (2001). Adaptív evolúciós megőrzés: a védelmi egységek meghatározásának egységes koncepciója felé. Mol. Ecol. 10, 2741–2752. doi: 10.1046 / j. 1365-294x.2001.t01-1-01411.x
PubMed Abstract / CrossRef Full Text / Google Scholar
Funk, W. C., Mckay, J. K., Hohenlohe, P. A., és Allendorf, F. W. (2012). A genomika hasznosítása a természetvédelmi egységek kijelölésére. Trendek Ecol. Evol. 27, 489–496. doi: 10.1016 / j.fa.2012.05.012
P>PubMed Abstract | CrossRef Full Text | Google Scholar
Garnett, S. T., and Christidis, L. (2017). A taxonómia anarchia akadályozza a megőrzést. Természet 546, 25-27. doi: 10.1038/546025a
P>PubMed Abstract | CrossRef teljes szöveg | Google Scholar
Gippoliti, S., Cotterill, F. P. D., Zinner, D., and Groves, C. P. (2018). A rendszertani tehetetlenség hatásai az afrikai patások sokféleségének megőrzésére: áttekintés. Biol. Rev.93, 115-130. doi: 10.1111 / brv.12335
P > PubMed Abstract / CrossRef teljes szöveg | Google Scholar
Grant, V. (1981). Növényi Speciáció. New York, NY: Columbia University Press.
P > PubMed Abstract / Google Scholar
Hamilton, C. W., and Reichard, S. H. (1992). Jelenlegi gyakorlat a használata alfajok, fajták és forma a besorolás a vadon élő növények. Taxon 41, 485-498. doi: 10.2307/1222819
CrossRef teljes szöveg/Google Scholar
Harvey, M. S., Rix, M. G., Framenau, V. W., Hamilton, Z. R., Johnson, M. S., Teale, R. J., et al. (2011). Az ártatlanok védelme: a rövid hatótávolságú endemikus taxonok tanulmányozása fokozza a védelmi eredményeket. Fordított. Syst. 25, 1–10. doi: 10.1071/IS11011
CrossRef teljes szöveg | Google Scholar
Hé, J., Waples, R. S., Arnold, M. L., Butlin, R. K., and Harrison, R. G. (2003). A fajok biológia és természetvédelem terén tapasztalható bizonytalanságának megértése és szembeállítása. Trendek Ecol. Evol. 18, 597–603. doi: 10.1016 / j.fa.2003.08.014
CrossRef Full Text / Google Scholar
Hoban, S. M., Hauffe, H. C., Perez-Espona, S., Arntzen, J. W., Bertorelle, G., Bryja, J., et al. (2013). A genetikai sokféleség előtérbe helyezése a természetvédelmi politika és a gazdálkodás terén. Conserv. Genet. Resour. 5, 593–598. doi: 10.1007 / s12686-013-9859-y
CrossRef Full Text / Google Scholar
Isaac, N. J., Mallet, J., and Mace, G. M. (2004). Taxonómiai infláció: hatása a makroökológiára és a természetvédelemre. Trendek Ecol. Evol. 19, 464–469. doi: 10.1016 / j.fa.2004.06.004
P>PubMed Abstract / CrossRef Full Text / Google Scholar
Isaac, N. J., Turvey, S. T., Collen, B., Waterman, C., and Baillie, J. E. (2007). Emlősök a szélén: természetvédelmi prioritások alapján fenyegetés és törzsfejlődés. PLoS ONE 2: e296. doi: 10.1371 / folyóirat.pone.0000296
P > PubMed Abstract / CrossRef teljes szöveg | Google Scholar
Jones, M. R., and Good, J. M. (2016). Célzott befogás az evolúciós és ökológiai genomikában. Mol. Ecol. 25, 185–202. doi: 10.1111 / mec.13304
P > PubMed Abstract / CrossRef Full Text / Google Scholar
Jorger, K. M., and Schrödl, M. (2013). Hogyan lehet leírni egy rejtélyes fajt? A molekuláris taxonómia gyakorlati kihívásai. Elöl. Zool. 10:59. doi: 10.1186/1742-9994-10-59
PubMed Abstract / CrossRef Full Text / Google Scholar
laikre, L., Allendorf, F., Aroner, L. C., Baker, S., Gregovich, D. P., Hansen, M. M., et al. (2010). A genetikai sokféleség elhanyagolása a biológiai sokféleségről szóló egyezmény végrehajtásában. Conserv. Biol. 24, 86–88. doi: 10.1111 / j. 1523-1739.2009. 01425.x
PubMed Abstract / CrossRef Full Text / Google Scholar
Laikre, L., Palm, S., and Ryman, N. (2005). A halak genetikai populációs szerkezete: a part menti övezetek kezelésére gyakorolt hatások. Ambio 34,111–119. doi: 10.1579/0044-7447-34.2.111
PubMed Abstract / CrossRef Full Text / Google Scholar
Laity, T., Laffan, S. W., González-Orozco, C. E., Faith, D. P., Rosauer, D. F., Byrne, M., et al. (2015). Phylodiversity, hogy tájékoztassa a védelmi politika: egy ausztrál példa. Sci. Teljes Környezet. 534, 131–143. doi: 10.1016 / j. scitotenv.2015.04.113 P>PubMed Abstract | CrossRef Full Text/Google Scholar
Leache, A. D., and Oaks, J. R. (2017). Az egyetlen nukleotid polimorfizmus (SNP) hasznossága. adatok filogenetika. Ann. Rev. Ecol. Syst. 48, 69–84. doi: 10.1146/annurev-ecolsys-110316-022645
CrossRef Full Text | Google Scholar
Llorens, T. M., Macdonald, B., McArthur, S., Coates, D. J., and Byrne, M. (2015). Disjunct, highly divergent genetic lineages within two rare Eremophila (Scrophulariaceae: Myoporeae). species in a biodiversity hotspot: implications for taxonomy and conservation. Biol. J. Linn. Soc. 177, 96–111. doi: 10.1111/boj.12228
CrossRef Full Text | Google Scholar
Love Stowell, S. M., Pinzone, C. A., and Martin, A. P. (2017). A genetikai sokféleség aktív beavatkozásainak akadályainak leküzdése. Biodiverzumok. Conserv. 26, 1753–1765. doi: 10.1007 / s10531-017-1330-z
CrossRef Full Text / Google Scholar
Mace, G. M. (2004). A taxonómia szerepe a fajok megőrzésében. Philos. Trans. R. Soc. B: Biol. Sci. 359, 711–719. doi: 10.1098 / rstb.2003.1454
P> PubMed Abstract | CrossRef teljes szöveg/Google Scholar
Mallet, J. (2007). Hibrid speciáció. Természet 446, 279-283. doi: 10.1038 / nature05706
P > PubMed Abstract / CrossRef teljes szöveg / Google Scholar
Mayr, E. (1942). Szisztematika és a fajok eredete. New York, NY: Columbia University Press.
Google Scholar
Mee, J. A., Bernatchez, L., Reist, J. D., Rogers, S. M., and Taylor, E. B. (2015). Az intraspecifikus megőrzési prioritások kijelölhető egységeinek azonosítása: hierarchikus megközelítés a fehérhal-tó fajkomplexumára (Coregonus spp.). Evol. Appl. 8, 423–441. doi: 10.1111 / eva.12247
PubMed Abstract | CrossRef Full Text | Google Scholar
Millar, M. A., and Byrne, M. (2013). Cryptic divergent lineages of Pultenaea pauciflora M.B. Scott (Fabaceae: Mirbelieae) exhibit different evolutionary history. Biol. J. Linn. Soc. 108, 871–881. doi: 10.1111/bij.12002
CrossRef Full Text | Google Scholar
Moritz, C. (1994). Defining evolutionarily significant units for conservation. Trends Ecol. Evol. 9, 373–375. doi: 10.1016/0169-5347(94)90057-4
CrossRef Full Text | PubMed Abstract
Moritz, C. (1999). Védelmi egységek és transzlokációk: stratégiák az evolúciós folyamatok megőrzésére. Hereditas 130, 217-228. doi: 10.1111 / j. 1601-5223.1999.00217.x
CrossRef Full Text / Google Scholar
Moritz, C. (2002). Stratégiák a biológiai sokféleség és az azt fenntartó evolúciós folyamatok védelmére. Syst. Biol. 51, 238–254. doi: 10.1080 / 10635150252899752
P>PubMed Abstract | CrossRef teljes szöveg / Google Scholar
Moritz, C., and Potter, S. (2013). Az evolúciós perspektíva fontossága a természetvédelmi politika tervezésében. Mol. Ecol. 22, 5969–5971. doi: 10.1111 / mec.12565
P > PubMed Abstract / CrossRef teljes szöveg | Google Scholar
Moritz, C., Pratt, R. C., Bank, S., Bourke, G., Bragg, J. G., Doughty, P., et al. (2018). Az Ausztrál gyíkok (Gehyra) egyik fajkomplexumában a rejtélyes leszármazási sokféleség, a testméret eltérése és a szimpátia. Evolúció 27, 54-66. doi: 10.1111 / evo.13380
CrossRef Full Text / Google Scholar
Morrison, W. R., Lohr, J. L., Duchen, P., Wilches, R., Trujillo, D., Mair, M., et al. (2009). A rendszertani változás hatása a megőrzésre: Megöli, meg tudja menteni, vagy csak irreleváns? Biol. Hátrányok. 142, 3201–3206. doi: 10.1016 / j. biocon.2009.07.019
CrossRef Full Text | Google Scholar
Murphy, N. P., Adams, M., Guzik, M. T., and Austin, A. D. (2013). Rendkívüli mikro-endemizmus az Ausztrál sivatagi tavaszi amphipodákban. Mol. Filogenet. Evol. 66, 645–653. doi: 10.1016 / j.ympev.2012.10.013
P>PubMed Abstract | CrossRef Full Text | Google Scholar
Murphy, N. P., King, R. A., and Delean, S. (2015). Fajok, esu-k vagy populációk? Az Ausztrál sivatagi tavaszi amphipodák morfológiailag rejtélyes sokféleségének meghatározása és leírása. Fordított. Syst. 29, 457–467. doi: 10.1071/IS14036
CrossRef teljes szöveg / Google Scholar
Nater, A., Mattle-Greminger, M. P., Nurcahyo, A., Nowak, M. G., De Manual, M., Desai, T., et al. (2017). Morphometric, behavioral, and genomic evidence for a new orangutan species. Curr. Cell Biol. 27, 3487–3498. doi: 10.1016 / j.cub.2017.09.047
CrossRef teljes szöveg | Google Scholar
Nicotra, A. B., Chong, C., Bragg, J. G., Ong, C. R., Aitken, N. C., Chuah, A., et al. (2016). Populáció és filogenomikus bomlás Genotipizálással-szekvenálással Ausztrál Pelargoniumban. Mol. Ecol. 25, 2000–2014. doi: 10.1111 / mec.13584
P > PubMed Abstract / CrossRef teljes szöveg | Google Scholar
Nistelberger, H., Byrne, M., Coates, D., and Roberts, J. D. (2014a). Erős filogeográfiai szerkezet egy millipede jelzi pleisztocén plébánia populációk között sávos vas képződmények félszáraz Ausztráliában. PLoS ONE 9: e0093038. doi: 10.1371 / folyóirat.pone.0093038
P > PubMed Abstract / CrossRef teljes szöveg | Google Scholar
Nistelberger, H., Gibson, N., Macdonald, B., Tapper, S.-L., and Byrne, M. (2014B). Phylogeographic bizonyíték két mesic refugia egy biodiverzitás hotspot. Öröklődés 113, 454-463. doi: 10.1038 / hdy.2014.46
P > PubMed Abstract / CrossRef Full Text / Google Scholar
Ogilvie, H. A., Heled, J., Xie, D., and Drummond, A. J. (2016). A beast számítási teljesítménye és statisztikai pontossága, valamint más módszerekkel történő összehasonlítás. Syst. Biol. 65, 381–396. doi: 10.1093 / sysbio / syv118
P > PubMed Abstract / CrossRef Full Text / Google Scholar
Oliver, P. M., Keogh, J. S., and Moritx, C. (2015). Az evolúciós sokféleség katalogizálásának és megértésének új megközelítései: az Ausztrál herpetológia perspektívája. Aust. J. Zool. 62, 417–430. doi: 10.1071/ZO14091
CrossRef Full Text/Google Scholar
Oliver, P. M., Laver, R. J., Demello Martins, F., Pratt, R. C., Hunjan, S., and Moritz, C. (2017). A gerinces endemizmus és az evolúciós refugium új hotspotja a trópusi Ausztráliában. Búvár. Distrib. 23, 53–66. doi: 10.1111 / ddi.12506
CrossRef teljes szöveg | Google Scholar
Oliver, P. M., and Lee, M. S. Y. (2010). A botanikai és zoológiai kódok akadályozzák a biológiai sokféleség kutatását azáltal, hogy megakadályozzák a meg nem nevezett új fajok közzétételét. Taxon 59, 1201-1205. doi: 10.2307/20773990
CrossRef Full Text/Google Scholar
Padial, J., Miralles, A., De la Riva, I., and Vences, M. (2010). A taxonómia integratív jövője. Elöl. Zool. 7:16. doi: 10.1186/1742-9994-7-16
PubMed Abstract / CrossRef Full Text / Google Scholar
Palsbøll, P. J., Bérubé, M., and Allendorf, F. W. (2007). A kezelési egységek azonosítása populációs genetikai adatok felhasználásával. Trendek Ecol. Evol. 22, 11–16. doi: 10.1016 / j.fa.2006.09.003
P>PubMed Abstract | CrossRef Full Text | Google Scholar
Patton, J. L., And Smith, M. F. (1994). Paraphyly, polyphyly, és a természet fajhatárokat Pocket Gophers (nemzetség Thomomys). Syst. Biol. 43, 11–26. doi: 10.1093 / sysbio / 43.1.11
CrossRef Full Text / Google Scholar
Pierson, J. C., Coates, D. J., Oostermeijer, J. G. B., Beissinger, S. R., Bragg, J. G., Sunnucks, P., et al. (2016). Genetikai tényezők a veszélyeztetett fajok helyreállítási terveiben három kontinensen. Elöl. Ecol. Environ. 14, 433–440. doi: 10.1002 / díj.1323
CrossRef Full Text / Google Scholar
Potter, S., Bragg, J. G., Peter, B. M., Bi, K., and Moritz, C. (2016). Phylogenomics at the tips: inferring lineages and their demography history in a tropical lizard, Carlia amax. Mol. Ecol. 25, 1367–1380. doi: 10.1111 / mec.13546
P>PubMed Abstract | CrossRef Full Text | Google Scholar
Potter, S., Xue, A. T., Bragg, J. G., Rosauer, D. F., Roycroft, E. J., and Moritz, C. (2018). A pleisztocén éghajlati változások diverzifikálják a trópusi szavannát. Mol. Ecol. 27, 520–532. doi: 10.1111 / mec.14441 / full
P > PubMed Abstract / CrossRef Full Text / Google Scholar
Rabosky, D. L., Hutchinson, M. N., Donnellan, S. C., Talaba, A. L., and Lovette, I. J. (2014). A fajok határainak filogenetikai szétszerelése az Ausztrál skinks (Scincidae: Ctenotus) széles körben elterjedt csoportjában. Mol. Filogenet. Evol. 77, 71–82. doi: 10.1016 / j.ympev.2014.03.026
P > PubMed Abstract / CrossRef teljes szöveg | Google Scholar
Ralls, K., Ballou, J. D., Dudash, M. R., Eldridge, M. D. B., Fenster, C. B., Lacy, R. C., et al. (2018). Paradigmaváltásra szólít fel a töredezett populációk genetikai kezelésében. Conserv. Lett. 11, 1–6. doi: 10.1111 / conl.124126
CrossRef teljes szöveg | Google Scholar
Rannala, B. (2015). A fajok elhatárolásának művészete és tudománya. Curr. Zool. 61, 846–853. doi: 1093.10.10./czoolo/61.5.846
CrossRef teljes szöveg/Google Scholar
Riedel, A., Sagata, K., Suhardjono, Y. R., Tänzler, R., and Balke, M. (2013). Integratív taxonómia a gyors pályán – a biológiai sokféleség kutatásának fenntarthatósága felé. Elöl. Zool. 10:15. doi: 10.1186/1742-9994-10-15
P>PubMed Abstract | CrossRef teljes szöveg | Google Scholar
Rieseberg, L. H., and Willis, J. H. (2007). Növényi speciáció. Tudomány 317, 910-914. doi: 10.1126 / tudomány.1137729
P > PubMed Abstract / CrossRef teljes szöveg / Google Scholar
Rosauer, D. F., Blom, M. P. K., Bourke, G., Catalano, S., Donnellan, S., Mulder, E., et al. (2016). Phylogeography, hotspots and conservation priorities: egy példa a felső végén Ausztrália. Biol. Hátrányok. 204, 83–93. doi: 10.1016 / j. biocon.2016.002
CrossRef teljes szöveg | Google Scholar
Rosauer, D. F., Byrne, M., Blom, M. P. K., Coates, D. J., Donnellan, S., Doughty, P., et al. (2018). Valós természetvédelmi tervezés evolúciós sokféleség a Kimberley, Ausztrália, sidesteps bizonytalan taxonómia Conserv. Lett. 11:e12438. doi: 10.1111 / conl.12438
CrossRef Full Text / Google Scholar
Rosenblum, E. B., Sarver, B. A. J., Brown, J. W., Des Roves, S., Hardwick, K. M., Hether, T. D., et al. (2012). A Goldilocks találkozik a Santa Rosalia-val: egy átmeneti speciációs modell magyarázza a diverzifikáció mintáit az időbeli skálákon. Evol. Biol. 39, 255–261. doi: 10.1007 / s11692-012-9171-x
PubMed Abstract / CrossRef teljes szöveg / Google Scholar
Roux, C., Fraïsse, C., Romiguier, J., Anciaux, Y., Galtier, N., and Bierne, N. (2016). A genomikus divergencia folytonossága mentén a speciáció szürke zónájára világít. PLoS Biol. 14:e2000234.hu itthon doi: 10.1371 / folyóirat.pbio.2000234
P > PubMed Abstract / CrossRef teljes szöveg | Google Scholar
Ryder, O. A. (1986). Fajmegőrzés és szisztematika-az alfajok dilemmája. Trendek Ecol. Evol. 1, 9–10. doi: 10.1016/0169-5347 (86)90059-5
CrossRef teljes szöveg | Google Scholar
Sampson, J. F., Byrne, M., Yates, C. J., Gibson, N., Thavornkanlapachai, R., Stankowski, S., et al. (2014). A kortárs pollen által közvetített gén bevándorlás tükrözi egy ritka cserje történelmi elszigeteltségét egy töredezett tájban. Öröklődés 112, 172-181. doi: 10.1038 / hdy.2013.89
P > PubMed Abstract / CrossRef Full Text / Google Scholar
Santamaría, L., and Méndez, P. F. (2012). A biológiai sokféleséggel kapcsolatos politika fejlődése: a jelenlegi hiányosságok és a jövőbeli igények. Evol. Appl. 5, 202–218. doi: 10.1111 / j. 1752-4571.2011.00229.x
P > PubMed Abstract / CrossRef Full Text / Google Scholar
Schlick-Steiner, B. C., Steiner, F. M., Seifert, B., Stauffer, C., Christian, E., and Crozier, R. H. (2010). Integratív taxonómia: többforrásos megközelítés a biológiai sokféleség feltárására. Annu. Rev. Entomol. 55, 421–438. doi: 10.1146/annurev-ento-112408-085432
P>PubMed Abstract | CrossRef teljes szöveg | Google Scholar
Seehausen, O, Takimoto, G., Roy, D., and Jokela, J. (2008). Speciation fordított biodiverzitás dinamika hibridizáció változó környezetben. Mol Ecol. 17, 30–44. doi: 10.1111 / j. 1365-294x.2007.03529.x
PubMed Abstract / CrossRef Full Text / Google Scholar
Shepherd, K. A., Thiele, K. R., Sampson, J., Coates, D. J., and Byrne, M. (2015). Egy új, ritka faj Atriplex (Chenopodiaceae), amely két, genetikailag különböző, de morfológiailag rejtélyes lakosság száraz Nyugat-Ausztrália: következmények taxonómia, illetve megőrzésére. Aust. Syst. Bot. 28, 234–245. doi: 10.1071 / SB15029
CrossRef teljes szöveg | Google Scholar
Singhal, S., and Bi, K. (2017). A történelem tisztítja a zűrzavarokat: az idő hatása a divergencia és az introgresszió vezetésére egy trópusi varrási zónában. Evolúció 71, 1888-1899. doi: 10.1111 / evo.13278
P > PubMed Abstract / CrossRef teljes szöveg | Google Scholar
Singhal, S., Hoskin, C., Couper, P., Potter, S., and Moritz, C. (2018). A rejtélyes Fajok megoldásának kerete: az Ausztrál nedves trópusok gyíkjainak esettanulmánya. Syst. Biol. . doi: 10.1093 / sysbio / syy
P > PubMed Abstract / CrossRef Full Text / Google Scholar
Singhal, S., and Moritz, C. (2013). A filogeográfiai vonalak közötti reprodukciós izoláció divergenciával skálázik. Proc. Roy. Soc B 280: 20132246. doi: 10.1098 / rspb.2013.2246026
P>PubMed Abstract / CrossRef Full Text / Google Scholar
Steane, D. A., Potts, B. M., McLean, E., Collins, L., Prober, S. M., Stock, W. D., et al. (2015). A genomra kiterjedő vizsgálatok feltárják a rejtélyes populáció szerkezetét egy száraz, adaptált eukaliptuszban. Fa Genet. Genomes 11:33. doi: 10.1007 / s11295-015-0864-z
CrossRef Full Text / Google Scholar
Struck, T. H., Feder, J. L., Bendiksby, M., Birkeland, S., Cerca, J., Gusarov, V. I., et al. (2017). A rejtélyes fajokban rejtett evolúciós folyamatok megtalálása. Trendek Ecol. Evol. 33, 153–163. doi: 10.1016 / j.fa.2017.11.007 P>PubMed Abstract | CrossRef Full Text/Google Scholar
Sukumaran, J., and Knowles, L. L. (2017). A többnemzetiségű coalescent határolja szerkezet nem faj. Proc. Nat. Aca. Sci. U. S. A. 114, 1607-1612. doi: 10.1073 / pnas.1607921114
P > PubMed Abstract / CrossRef Full Text / Google Scholar
Swarts, N. D., Clements, M. A., Bower, C. C., and Miller, J. T. (2014). A természetvédelmi egységek meghatározása morfológiailag hasonló, szexuálisan megtévesztő, erősen veszélyeztetett orchideák komplexumában. Biol. Hátrányok. 174, 55–64. doi: 10.1016 / j. biocon.2014.03.017
CrossRef teljes szöveg/Google Scholar
Tautz, D., Arctander, P., Minelli, A., Thomas, R. H., and Vogler, A. P. (2003). A DNS-taxonómia iránti kérelem. Trendek Ecol. Evol. 18, 70–74. doi: 10.1016 / S0169-5347 (02)00041-1
CrossRef teljes szöveg | Google Scholar
Taylor, H. R., Dussex, N., and van Heezik, Y. (2017). A természetvédelmi genetikai különbség áthidalása azáltal, hogy azonosítja a természetvédelmi szakemberek végrehajtásának akadályait. Glob. Ecol. Conserv. 10, 231–242. doi: 10.1016 / j. gecco.2017.04.001
CrossRef teljes szöveg | Google Scholar
Thomson, S. A., Pyle, R. L., Ahyong, S. T., Alonso-Zarazaga, M., Ammirati, J., Araya, J. F., et al. (2018). A tudományon alapuló taxonómia szükséges a globális megőrzéshez. PLoS Biol. 16: e2005075. doi: 10.1371 / folyóirat.pbio.2005075
P > PubMed Abstract / CrossRef Full Text / Google Scholar
Vieites, D. R., Wollenberg, K. C., Andreonec, F., Köhler, J., Glaw, F., And Vences, M. (2009). Madagaszkár biodiverzitásának hatalmas alábecsülése, amelyet egy integratív kétéltű leltár bizonyít. Proc. NAT. Acad. Sci. U. S. A. 106, 8267-8272. doi: 10.1073 / pnas.0810821106
CrossRef teljes szöveg
Vogler, A. P., and de Salle, R. (1994). A védelmi menedzsment egységeinek diagnosztizálása. Hátrányok. Biol. 8, 354–363. doi: 10.1046 / j. 1523-1739.1994. 08020354.x
CrossRef Full Text / Google Scholar
Waples, R. S. (1991). Csendes-óceáni lazac, Oncorhynchus spp., valamint a “faj” meghatározása a veszélyeztetett fajokról szóló törvény alapján. Mar. Hal. 53. perc: 11-22.
Google Scholar
Waples, R. S. (1995). “Evolúciósan jelentős egységek és a biológiai sokféleség megőrzése a veszélyeztetett fajokról szóló törvény szerint”, az evolúcióban és a vízi ökoszisztémában: a populáció megőrzésének egyedi egységeinek meghatározása, eds J. L. Nielsen (Bethesda, MD:American Fisheries Society), 8-27.
Google Scholar
Waples, R. S., Gustafson, R. G., Weitkamp, L. A., Myers, J. M., Jjonson, O. W., Busby, P. J., et al. (2001). A lazac sokféleségének jellemzése A Csendes-óceán északnyugati részéről. J. Fish Biol. 59, 1–41. doi: 10.1111 / j. 1095-8649. 2001.tb01376.x
CrossRef Full Text / Google Scholar
Waples, R. S., Nammack, M., Cochrane, J. F., and Hutchings, J. A. (2013). Két cselekmény története: veszélyeztetett fajok listája Kanadában és az Egyesült Államokban. BioScience 63, 723-734. doi: 10.1093 / bioscience / 63.9.723
CrossRef Full Text / Google Scholar
Waples, R. S., Punt, A. E., and Cope, J. M. (2008). A genetikai adatok integrálása a tengeri erőforrások kezelésébe: hogyan tehetjük ezt jobban? Hal 9, 423-449. doi: 10.1111 / j. 1467-2979.2008.00303.x
CrossRef Full Text / Google Scholar
Wege, J. A., Thiele, K. R., Shepherd, K. A., Butcher, R., Macfarlane, T. D., and Coates, D. J. (2015). Stratégiai taxonómia egy biodiverzális tájban: újszerű megközelítés a ritka és kevéssé ismert flóra természetvédelmi eredményeinek maximalizálására. Biodiverzumok. Conserv. 24, 17–32. doi: 10.1007 / s10531-014-0785-4
CrossRef Full Text / Google Scholar
Whitlock, R. (2014). Az adaptív és semleges genetikai sokféleség, valamint az ökológiai struktúra és működés közötti kapcsolatok: meta-analízis. J. Ecol. 102, 857–872. doi: 10.1111/1365-2745.12240
P>PubMed Abstract | CrossRef teljes szöveg | Google Scholar
Wiley, E. O. (1978). Az evolúciós faj fogalmát újraértékelték. Syst. Biol. 27, 17–26. doi: 10.2307 / 2412809
CrossRef teljes szöveg | Google Scholar
Wilson, E. O. (2017). A biodiverzitás kutatásához több bakancsra van szükség a földön. Nat. Ecol. Evol. 1, 1590–1591. doi: 10.1038 / s41559-017-0360-y
PubMed Abstract / CrossRef Full Text / Google Scholar
Yang, Z. H., and Rannala, B. (2014). Nem irányított Fajok elhatárolása több lókuszból származó DNS-szekvencia-adatok felhasználásával. Mol. Biol. Evol. 31, 3125–3135. doi: 10.1093/molbev/msu279
P>PubMed Abstract | CrossRef teljes szöveg | Google Scholar
Yeates, D. K., Seago, A., Nelson, L., Cameron, S. L., Joseph, L., and Trueman, W. H. (2011). Integratív taxonómia, vagy iteratív taxonómia? Sys. Ent. 36, 209–217. doi: 10.1111/j.1365-3113.2010.00558.x
CrossRef Full Text | Google Scholar