VIVO Patofysiologia
insuliinisynteesi ja eritys
insuliini on pieni proteiini, jonka molekyylipaino on noin 6000 daltonia. Se koostuu kahdesta ketjusta, joita disulfidisidokset pitävät koossa. Oikealla olevassa kuvassa on naudan insuliinin molekyylimalli, jossa A-ketju on väriltään sininen ja suurempi B-ketju vihreä. Voit saada paremman arvostuksen insuliinin rakenteesta manipuloimalla tällaista mallia itse.
aminohappojärjestys säilyy selkärankaisilla hyvin, ja yhden nisäkkään insuliini on lähes varmasti biologisesti aktiivinen toisessa. Vielä nykyäänkin monia diabeetikoita hoidetaan sian haimasta uutetulla insuliinilla.
insuliinin biosynteesi
insuliinia syntetisoidaan merkittäviä määriä vain haiman beetasoluissa. MRNA-insuliini muunnetaan yksiketjuiseksi esiasteeksi, jota kutsutaan preproinsuliiniksi, ja sen signaalipeptidin poistaminen endoplasmaiseen verkkokalvoon asetettaessa tuottaa proinsuliinia.
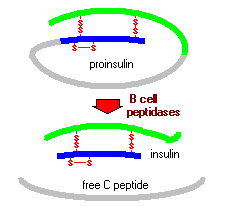 proinsuliini koostuu kolmesta domeenista: aminoterminaalisesta B-ketjusta, karboksyyliterminaalisesta a-ketjusta ja keskellä olevasta yhdistävästä peptidistä, joka tunnetaan nimellä C-peptidi. Endoplasmaisessa retikulumissa proinsuliini altistuu useille spesifisille endopeptidaaseille, jotka valmistavat C-peptidin ja tuottavat näin kypsän insuliinin muodon. Insuliini ja vapaa C-peptidi pakataan Golgiin sekretorisiksi rakeiksi, jotka kerääntyvät sytoplasmaan.
proinsuliini koostuu kolmesta domeenista: aminoterminaalisesta B-ketjusta, karboksyyliterminaalisesta a-ketjusta ja keskellä olevasta yhdistävästä peptidistä, joka tunnetaan nimellä C-peptidi. Endoplasmaisessa retikulumissa proinsuliini altistuu useille spesifisille endopeptidaaseille, jotka valmistavat C-peptidin ja tuottavat näin kypsän insuliinin muodon. Insuliini ja vapaa C-peptidi pakataan Golgiin sekretorisiksi rakeiksi, jotka kerääntyvät sytoplasmaan.
kun beetasolua stimuloidaan asianmukaisesti, insuliini erittyy solusta eksosytoosilla ja diffundoituu saarten kapillaarivereen. C-peptidi erittyy myös vereen, mutta sillä ei ole tunnettua biologista aktiivisuutta.
Insuliininerityksen kontrollointi
insuliini erittyy pääasiassa kohonneiden veren glukoosipitoisuuksien seurauksena. Tämä on järkevää, koska insuliini ”vastaa” glukoosin pääsyn helpottamisesta soluihin. Jotkut hermoärsykkeet (esim. ruoan näkö ja maku) ja muiden polttoainemolekyylien, kuten aminohappojen ja rasvahappojen, kohonneet pitoisuudet veressä edistävät myös insuliinin eritystä.
käsityksemme insuliinin erityksen taustalla olevista mekanismeista on edelleen jokseenkin hajanainen. Tämän prosessin tietyt piirteet on kuitenkin selvästi ja toistuvasti osoitettu, ja tuloksena on seuraava malli:
- glukoosi kulkeutuu beetasoluun fasilitoidun diffuusion kautta glukoosikuljettajan kautta; kohonneet glukoosipitoisuudet solunulkoisessa nesteessä johtavat kohonneisiin glukoosipitoisuuksiin beetasolussa.
- kohonneet glukoosipitoisuudet beetasolussa johtavat lopulta kalvon depolarisaatioon ja solunulkoisen kalsiumin virtaukseen. Tästä johtuvan solunsisäisen kalsiumin lisääntymisen ajatellaan olevan yksi tärkeimmistä laukaisevista tekijöistä insuliinia sisältävien sekretoristen rakeiden eksosytoosille. Mekanismit, joilla kohonneet glukoositasot beetasolussa aiheuttavat depolarisaatiota, eivät ole selvästi selvillä, vaan ne näyttävät johtuvan glukoosin ja muiden polttoainemolekyylien aineenvaihdunnasta solussa, ehkä aistittuna ATP:ADP-suhteen muutoksena ja muuntautuneena kalvonjohtavuuden muutoksiksi.
- glukoosipitoisuuden nousu beetasoluissa näyttää myös aktivoivan kalsiumista riippumattomia reittejä, jotka osallistuvat insuliinin eritykseen.
 insuliinin vapautumisen stimulaatiota havaitaan helposti kokonaisilla eläimillä tai ihmisillä. Normaali paastoverenglukoosipitoisuus ihmisillä ja useimmilla nisäkkäillä on 80-90 mg / 100 ml, mikä liittyy hyvin alhaiseen insuliinineritykseen.
insuliinin vapautumisen stimulaatiota havaitaan helposti kokonaisilla eläimillä tai ihmisillä. Normaali paastoverenglukoosipitoisuus ihmisillä ja useimmilla nisäkkäillä on 80-90 mg / 100 ml, mikä liittyy hyvin alhaiseen insuliinineritykseen.
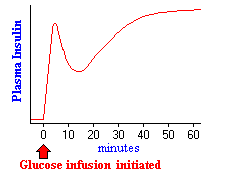
oikealla oleva kuva kuvaa vaikutuksia insuliinin eritykseen, kun infusoidaan riittävästi glukoosia pitämään veren pitoisuuden kaksi-kolme kertaa paastotason suuruisena tunnin ajan. Lähes välittömästi infuusion alkamisen jälkeen plasman insuliinipitoisuudet nousevat dramaattisesti. Tämä alkuvaiheen lisääntyminen johtuu esimuotoutuneen insuliinin erityksestä, joka loppuu pian merkittävästi. Insuliinin sekundäärinen nousu heijastaa sitä huomattavaa määrää uutta syntetisoitua insuliinia, joka vapautuu välittömästi. On selvää, että kohonnut glukoosi ei ainoastaan simuloi insuliinin eritystä, vaan myös insuliinigeenin transkriptiota ja sen mRNA: n translaatiota.
syventävät ja täydentävät aiheet
- insuliinin rakenne