la începutul anilor 1960, biologii au început să investigheze modul în care proteinele la diferite specii evoluează la nivel de secvență (Zuckerkandl 1962, margoliash 1963, Doolittle & blomb inktck 1964). Proteinele analizate au inclus hemoglobina (Zuckerkandl & Pauling 1962), citocromul c (Margoliash 1963) și fibrinopeptidele (Doolittle& Blomb 1964). Aceste investigații timpurii au condus la o descoperire remarcabilă: se pare că numărul diferențelor dintre secvențele de proteine ale diferitelor specii a fost aproximativ proporțional cu timpul de la divergența speciilor (Figura 1).
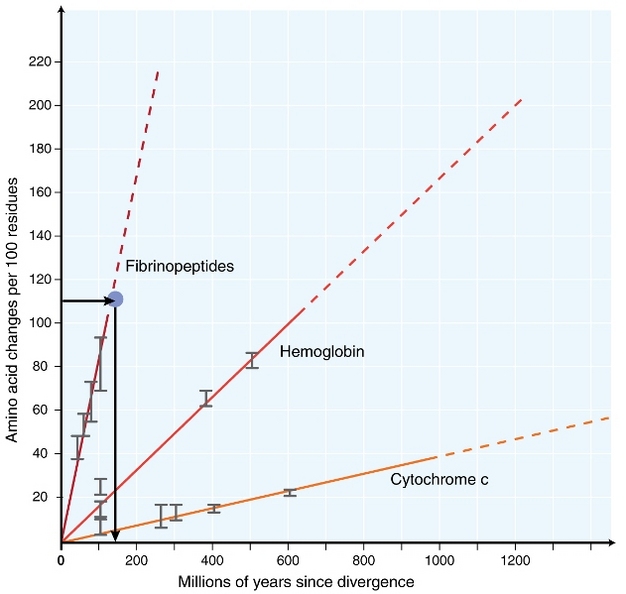
Zuckerkandl& Pauling (1965) a comparat acumularea constantă de substituții de aminoacizi în timp cu „căpușe” regulate ale ceasurilor și a afirmat că „poate exista un ceas evolutiv molecular”. Astfel, termenul ‘ceas molecular’ a fost inițial inventat pentru a descrie modificările aminoacizilor care apar proporțional cu timpul de la divergența speciilor.
de la prima utilizare, termenul ‘ceas molecular’ a fost folosit în multe contexte diferite. În zilele noastre, este adesea folosit pentru a se referi pur și simplu la numărul de modificări sau substituții acumulate în secvența de ADN sau proteine dintr-o anumită linie. Numărul de substituții pe o unitate de timp definită poate fi descris ca ‘rata’ ceasului molecular, care în acest context este echivalent cu ‘rata evolutivă’. Rețineți că conotația inițială este încă valabilă în unele cazuri. De exemplu, atunci când oamenii afirmă ‘Am presupus un ceas molecular’ (în special în analizele filogenetice), înseamnă că s-a presupus că numărul de substituții apare constant în timp.
conceptul unui ceas molecular constant are implicații extraordinare pentru biologii evoluționiști. Dacă un ceas molecular constant, așa cum s-a propus inițial, ar exista cu adevărat, deducerea calendarului evenimentelor evolutive ar deveni o problemă destul de simplă (Figura 1). Cu toate acestea, a devenit foarte clar că substituțiile nu apar în mod constant în timp în diferite linii (Kumar 2005). Cu toate acestea, conceptul de ceas molecular a fost extrem de influent în domeniul evoluției moleculare. Una dintre cele mai importante idei inspirate de conceptul ceasului molecular este teoria neutră a evoluției moleculare.
ceasul Molecular și neutralitatea
când a apărut pentru prima dată ideea unui ceas molecular constant, s-a crezut că forța evolutivă predominantă care stă la baza substituțiilor de aminoacizi sau nucleotide a fost selecția naturală. Urmând această linie de gândire, un ceas molecular constant ar indica faptul că substituțiile adaptive la diferite specii apar constant în timp. Cu toate acestea, este greu de explicat modul în care substituțiile adaptive ar avea loc într-un mod atât de asemănător ceasului. Teoretic, soarta mutațiilor adaptive este determinată de mai mulți parametri evolutivi, cum ar fi puterea avantajului selectiv al acelei mutații, dimensiunea populației efective și ratele de mutație adaptivă (Kimura 1983). Acești parametri sunt susceptibili să difere între specii și chiar în cadrul unei specii, în funcție de mutații specifice și de interacțiunile lor cu mediile. în schimb, Kimura (1968, 1969) a propus că majoritatea schimbărilor la nivel molecular au consecințe funcționale reduse sau sunt ‘neutre’. Dacă o mutație nu are consecințe de fitness, soarta sa în populație este determinată complet de întâmplare aleatorie. Aceasta înseamnă că nu putem prezice dacă o mutație neutră specifică va fi în cele din urmă fixată în populație. Cu toate acestea, rata la care apar substituții neutre în populație poate fi prezisă deoarece depinde de un singur parametru, și anume rata mutației (Kimura 1968).
să ne imaginăm o populație cu N număr de indivizi haploizi. Dacă mutațiile neutre apar la rata u pe individ pe generație, numărul total de mutații dintr-o generație va fi de N ori u. deoarece toate aceste mutații noi sunt neutre, destinele lor sunt complet determinate întâmplător. Cu alte cuvinte, toate mutațiile au șanse egale de a ajunge la fixare (ceea ce duce la o ‘substituție’). Probabilitatea ca fiecare nouă mutație neutră să ajungă la fixare, având în vedere că a avut loc o substituție, este pur și simplu 1/N. Rata substituțiilor este calculată ca numărul de mutații noi din fiecare generație (Nu) înmulțit cu probabilitatea ca fiecare mutație nouă să atingă fixarea (1/N), care este egală cu u. cu alte cuvinte, pentru mutațiile neutre, rata de substituție este egală cu rata mutației! prin urmare, dacă majoritatea mutațiilor sunt neutre (așa cum se propune în teoria neutră) și dacă ratele de mutație sunt constante în timp, substituțiile ar trebui să apară constant și în timp. Ar trebui să observăm apoi rate regulate de substituții asemănătoare ceasului la nivel molecular. Kimura (1969) a considerat astfel observarea ceasului molecular relativ constant în secvențele de proteine ca un sprijin puternic pentru teoria neutră a evoluției moleculare.
testarea modului în care funcționează ceasul Molecular neutru
conform teoriei neutre, întrebarea dacă ratele de substituție sunt constante în timp sau nu este echivalentă cu dacă ratele de mutație neutre sunt constante în timp. Din acest motiv, multe studii ulterioare s-au concentrat pe analiza datelor din site-uri neutre pentru a determina dacă ratele de mutație neutre sunt într-adevăr constante în timp. Vom analiza pe scurt modul în care aceste studii sunt puse în aplicare, înainte de a discuta dezbateri teoretice asupra ceasurilor moleculare. Analizele ceasurilor moleculare proteice au continuat, de asemenea, dar dezbaterile din jurul variației ceasurilor moleculare proteice sunt foarte diferite de cele referitoare la ceasurile moleculare neutre și nu vor fi incluse în acest articol. Cititorii interesați ar trebui să consulte Gillespie (1991), Kumar (2005), Kim și Yi (2008) și Bedford și colab. (2008).
majoritatea analizelor empirice ale ceasurilor moleculare neutre se bazează pe teorema conform căreia ratele de mutație neutre pot fi deduse din ratele de substituție neutre (Kimura 1968, 1969). În practică, fiecare studiu definește un anumit tip de site-uri din genom ca site-uri neutre și compară ratele de substituție ale acestor site-uri între linii.
care site-uri din genom sunt cu adevărat neutre nu pot fi determinate complet, dar oamenii de știință au venit cu mai multe proxy-uri utile. Înainte de era secvențierii genomului, cele mai multe date de secvență disponibile erau cele din secvențele ADN care codifică proteinele. Studiile împart adesea secvențele ADN care codifică proteinele în două tipuri de site-uri (Wu & Li 1985). Primul tip de site-uri include cele pentru care orice modificare ar duce la substituții de aminoacizi, sau ‘site-uri nonsinonymous’. Al doilea tip de site-uri, ‘site-uri Sinonime’, include cele care codifică pozițiile ‘degenerate’ în tabelul codon, unde o modificare nu duce la o substituție de aminoacizi. De exemplu, TCT și TCC codifică Serina. Dacă se schimbă A treia poziție a acestor codoni, aceștia vor produce în continuare aceiași aminoacizi. Astfel de substituții ar fi mai puțin vizibile pentru selecția naturală. În consecință, ceasurile moleculare din locurile sinonime ar trebui să fie mai aproape de ceasul molecular neutru decât ceasurile nonsinonime.
pe măsură ce tehnicile de secvențiere au avansat, unele studii au folosit intronii ca proxy pentru siturile neutre, deoarece nu sunt încorporați în ARNm maturi și, prin urmare, sunt mai susceptibili de a fi neutri (Yi și colab. 2002). Secvențe de elemente transpozabile inactive care au fost inserate cu mult înainte de divergența speciilor au fost, de asemenea, adesea folosite (acestea sunt adesea denumite ‘repetiții ancestrale’, de exemplu, Thomas și colab., 2003). În cele din urmă, unele studii au folosit secvențe ADN necodificatoare (toate secvențele după îndepărtarea secvențelor ADN care codifică proteinele) extrase din aliniamentele genomului întreg pentru a testa ceasurile moleculare neutre (Elango și colab. 2006, Huttley și colab. 2007).
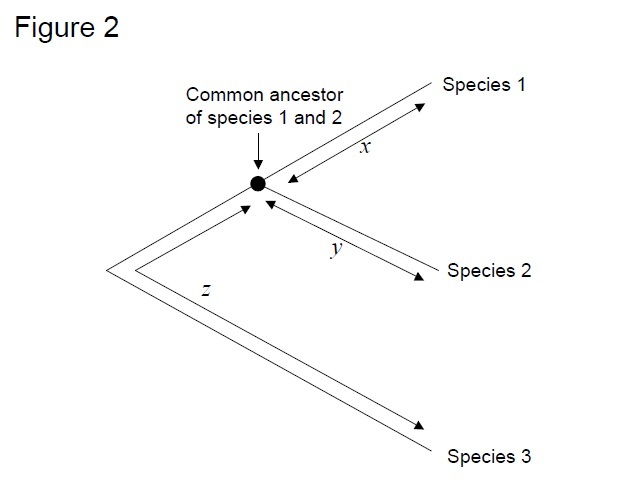
testul cel mai frecvent utilizat este așa-numitul”test de rată relativă”(Sarich& Wilson 1973). Inițial, ratele de substituție pe unitate de timp au fost estimate prin împărțirea numărului total de diferențe (substituții) între proteinele diferitelor specii la timpul de divergență, estimat din înregistrările fosile (Figura 1). Cu toate acestea, înregistrările fosile nu sunt disponibile pentru multe comparații și sunt asociate cu marje mari de eroare. Testul ratei relative depășește necesitatea înregistrărilor fosile (Figura 2). Atâta timp cât există o secvență outgroup la cele două linii de interes, putem determina dacă cele două ramuri urmează aceleași ceasuri moleculare sau diferite, fără cunoașterea timpului absolut de divergență (Figura 2).
care sunt factorii determinanți ai ceasurilor moleculare neutre?
aproape toate controversele din centrul dezbaterilor asupra ceasurilor moleculare neutre provin din întrebarea care sunt sursele majore de mutații. Această întrebare este direct relevantă pentru înțelegerea modelelor de mutație, care sunt sursa finală a schimbărilor evolutive și a bolilor genetice. Mai mult, înțelegerea modului în care ratele de mutație variază între linii și în cadrul genomilor este o întrebare fundamentală în genomica comparativă, care își propune să utilizeze comparații de secvențe pentru a identifica regiunile genomice care sunt importante din punct de vedere funcțional.
deci, ce determină ratele de mutație neutre? Unul dintre cei mai importanți contribuitori la ceasurile moleculare neutre este variația specifică liniei în timpii de generație. De la început, ideea unui ceas molecular neutru constant a fost percepută ca fiind în contradicție cu mecanismele moleculare ale mutației liniei germinale. S-a considerat mult timp că majoritatea mutațiilor apar din erori în replicarea ADN-ului în linii germinale (Haldane 1947, Muller 1954). Deoarece mutațiile apar atunci când ADN-ul germinal este reprodus pentru următoarea generație, acestea ar trebui să se acumuleze proporțional cu numărul de generații, mai degrabă decât cu timpul absolut. Prin urmare, dacă am compara numărul de substituții care s-au acumulat în două linii de la divergența lor, linia cu un timp de generație mai lung, care a suferit mai puține evenimente de replicare a ADN-ului, ar avea mai puține substituții în comparație cu linia cu timpul de generație mai scurt. În consecință, ceasul molecular ar trebui să funcționeze mai lent la speciile cu timpi de generație mai lungi. Această idee este denumită efectul generației-timp.de fapt, efectul timpului de generare a fost observat pentru prima dată în studiile primatelor, chiar înainte de dezbaterea privind ceasul molecular. Morris Goodman, care folosea metode imunologice pentru a investiga legătura dintre specii la acea vreme, a observat că rata la care unele proteine diferă pare să fie scăzută la maimuțe, în special la oameni, comparativ cu maimuțele din Lumea Veche (Goodman 1961, 1962, 1963). Acest efect este denumit ‘încetinirea ratei hominoide’. Deoarece hominoidele au timpi de generație mai lungi decât maimuțele din Lumea Veche, această observație poate fi explicată prin efectul timpului de generație.
Wu& Li (1985) au fost primii care au testat ipoteza efectului generație-timp folosind date de secvență ADN. Au folosit date de la 11 gene de primate și rozătoare. Deoarece primatele au un timp de generare mult mai lung decât rozătoarele, ceasul molecular ar trebui să fie mai rapid la rozătoare în comparație cu primatele. Într-adevăr, au descoperit că pentru site-urile sinonime, rozătoarele prezintă aproximativ de două ori rata de evoluție moleculară în comparație cu primatele (Wu & Li 1985). Cu toate acestea, pentru site-urile nesinonime, un astfel de efect nu a fost găsit. Cu alte cuvinte, ceasul molecular neutru, dar nu ceasul molecular al aminoacizilor, căpușează mai repede în linia rozătoarelor în comparație cu linia primatelor, ceea ce se potrivește bine cu ideea unui efect de generație-timp.
studiile ulterioare au oferit sprijin suplimentar încetinirii ratei hominoide (Li & Tanimura 1987, Bailey și colab. 1991) și diferența de rată dintre rozătoarele și primatele (Gu & Li 1992, Huttley și colab. 2007). Mai mult, o diferență de rată a fost observată la scări filogenetice și mai mici, în special la primate: de exemplu, ceasul molecular uman rulează mai lent decât ceasul molecular al cimpanzeului (Elango și colab. 2006); ratele la maimuțele din Lumea Nouă sunt mai rapide decât ratele la hominoizi și maimuțele din Lumea Veche (Steiper & Young 2006). Ratele diferite ale ceasurilor moleculare observate în aceste studii sunt în concordanță calitativă cu efectul de generare-timp.
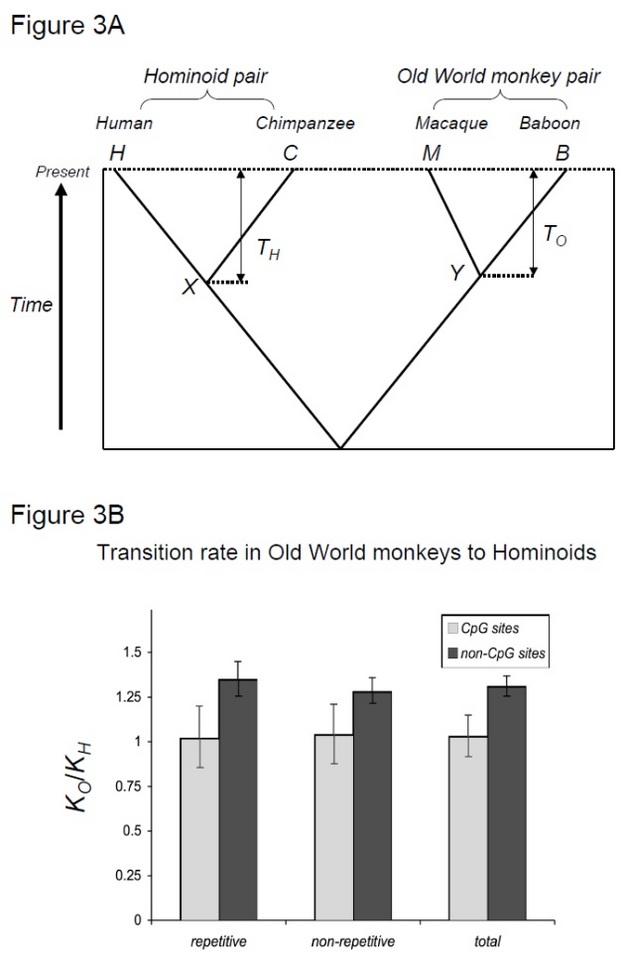
cu toate acestea, diferențele reale dintre linii nu sunt cantitativ în concordanță cu diferența în timpii de generație. De exemplu, Kumar & Subramanian (2002) a arătat că, deși diferența de timp de generație între primate și rozătoare este mult mai mare decât cea dintre oameni și maimuțele lumii vechi, diferențele observate în ceasurile moleculare sunt similare în aceste două comparații. Merită menționat faptul că Kumar & Subramanian (2002) a folosit filtre statistice specifice pentru a elimina datele care arătau modele de substituție „eterogene”, care ar fi putut provoca o părtinire către secvențe care evoluează lent (Yi și colab. 2002). Cu toate acestea, diferența dintre ceasurile moleculare ale primatelor și rozătoarelor pare mult mai mică decât cea propusă inițial de Wu & Li (1985). De exemplu, Huttley și colab. (2007) a analizat alinierea întregului genom al mai multor specii, inclusiv opossum, și a arătat că diferența de rată dintre descendențele eutheriene și descendența opossum (~30%) este mult mai mare decât diferența de rată dintre descendențele umane și șoareci (~14%). Aceste exemple demonstrează că gradul de diferențe în ceasurile moleculare variază semnificativ între diferite studii datorită diferențelor dintre seturile de date și metodele statistice. Acestea arată, de asemenea, că diferențele de rată dintre linii nu pot fi complet contabilizate doar de diferența dintre timpii de generație. În mod clar, există și alți contribuitori la ceasurile moleculare neutre. într-adevăr, trăsăturile istoriei vieții, altele decât timpurile de generație, par să co-varieze cu ceasurile moleculare. Martin& Palumbi (1993) a arătat că ceasurile moleculare ADN rulează mai repede la speciile cu dimensiuni mici ale corpului. Această observație a condus la ipoteza că ratele metabolice sunt factori determinanți importanți ai ceasurilor moleculare. O rată metabolică ridicată produce un număr mare de radicali mutageni de oxigen, ceea ce ar crește ratele de mutație (Rand 1994). Deoarece ratele metabolice și dimensiunea corpului variază în general în funcție de timpii de generație, a fost dificil să se distingă care dintre acestea constituie principalul determinant al ratelor moleculare de ceas. Tsantes & Steiper (2009) au propus, pe baza datelor de la primate, că vârsta la prima reproducere, mai degrabă decât dimensiunea corpului, este principalul determinant al ceasurilor moleculare. Deoarece vârsta la prima reproducere reflectă efectul timpului de generare, acest studiu susține ideea că efectul timpului de generare este principalul determinant al ceasului molecular. Cu toate acestea, acest studiu se bazează încă pe un număr limitat de linii (au fost utilizate patru perechi de specii). Astfel, distincția între dimensiunea corpului, efectul timpului de generare și ratele metabolice rămâne în continuare o problemă importantă în generalizarea și înțelegerea ceasurilor moleculare neutre. mai mult, importanța factorilor care nu variază în funcție de timpii de generație, cum ar fi metilarea ADN-ului, a fost din ce în ce mai apreciată. Metilarea ADN-ului este o modificare chimică a ADN-ului genomic găsit în diverși taxoni. În genomii animale, metilarea ADN-ului are loc aproape exclusiv la citozine urmate de guanine (așa-numitele CpG). Citozinele metilate, la rândul lor, tind să se mute rapid la timine din cauza instabilității chimice (Bird 1980). Într-adevăr, în genomul uman, mutațiile cauzate de metilarea ADN apar mai mult decât un ordin de mărime mai frecvent decât alte mutații (Nachman & Crowell 2000, Elango și colab. 2008). Deoarece mutațiile cauzate de metilarea ADN-ului apar în mare parte independent de replicarea ADN-ului, astfel de mutații pot urma Ceasuri moleculare diferite decât altele. Mai exact, în loc de dependența de generație-timp, mutațiile cauzate de metilarea ADN-ului pot urma un ceas molecular dependent de timp, care este similar cu ceea ce a fost propus inițial de Zuckerkandl & Pauling (Kim și colab. 2006)! pentru a testa această ipoteză, Kim și colab. (2006) a comparat divergența om-cimpanzeu cu divergența macac-babuin, două perechi de specii care împărtășesc timpi de divergență similari, dar cu timpi de generație diferiți (Steiper și colab. 2004). Perechea om-cimpanzeu (perechea hominoidă) are timpi de generație mult mai lungi în comparație cu perechea macac-babuin (perechea de maimuțe din Lumea Veche). Acest studiu a arătat că pentru site-urile non-CpG, perechea de maimuțe din Lumea Veche a acumulat cu aproximativ 30% mai multe substituții, ceea ce poate fi explicat prin efectul de încetinire a ratei hominoide menționat mai sus. În schimb, ceasurile moleculare din siturile CpG au arătat un număr similar de substituții în perechile de maimuțe hominoide și din Lumea Veche (Figura 3). Astfel, ceasurile moleculare dependente de timp și de generare dependente de timp coexistă în cadrul acelorași genomi. Presupunerea că un singur ceas molecular poate exista pentru o anumită linie nu mai este valabilă, deoarece forțele mutaționale predominante variază între regiunile genomice.
concluzii
conceptul de ceas molecular constant a fost inițial propus pe baza unei cantități limitate de date de secvență de proteine. Chiar dacă studiile ulterioare au arătat că o astfel de observație nu este un model general în aminoacizi, a avut o influență semnificativă asupra domeniului evoluției moleculare, în special asupra dezvoltării teoriei neutre a evoluției moleculare pentru datele secvenței ADN. Urmând teoria neutră, studiile s-au concentrat pe elucidarea tiparelor de variație a ratelor de mutație neutră. În ultimele decenii, am observat că ceasurile moleculare rulează la rate diferite între linii. În plus, gradul de variație poate varia în funcție de diferitele tipuri de date și de metodele statistice specifice utilizate. Efectul de generare-timp continuă să se mențină la un nivel calitativ, dar este insuficient pentru a explica variația cantitativă a ratelor de mutație neutră între linii. Trăsăturile istoriei vieții și mutațiile dependente de non-replicare, cum ar fi cele cauzate de metilarea ADN-ului, sunt, de asemenea, contribuitori importanți la ceasurile moleculare genomice. Într-adevăr, diferite tipuri de ceasuri moleculare sunt observate chiar și în cadrul unui genom, deoarece intrările mutaționale predominante variază între diferite regiuni genomice. Astfel, mai degrabă decât să presupunem un singur ceas molecular neutru pentru fiecare genom, studiile viitoare ar trebui să urmărească să dezvăluie variația ceasurilor moleculare neutre genomice, pentru a afla despre peisajele mutaționale genomice. Astfel de informații nu sunt utile doar pentru înțelegerea materiei prime care guvernează evoluția moleculară și boala genetică, ci constituie și o componentă critică care influențează analizele genomice comparative și funcționale pentru a identifica regiunile genomice funcționale.