plasticitatea musculară
mușchiul scheletic este cel mai mare țesut din corp. Este foarte plastic și poate suferi schimbări dramatice non-patologice atât în masă, cât și în forță, ca răspuns la o multitudine de influențe de mediu. În urma exercițiilor de rezistență sau a expunerii la steroizi anabolizanți, mușchii suferă hipertrofie și își măresc masa, aria secțiunii transversale și forța (Aagaard, 2004; Schiaffino și colab., 2013). Aceiași parametri pot scădea precipitat atunci când mușchii inițiază atrofie ca răspuns la oricare dintre gama de insulte care includ: înfometare, denervare, imobilizare, sepsis, cașexie canceroasă, insuficiență cardiacă, diabet, insuficiență renală, tulburare pulmonară obstructivă cronică și sindromul imunodeficienței dobândite (Schiaffino și colab., 2013).
consecințele atrofiei au implicații evidente pentru sănătate. Slăbiciunea musculară contribuie major atât la mortalitate, cât și la morbiditate și este asociată cu riscul crescut de toate cauzele de deces (Metter și colab., 2002). De fapt, reducerea atrofiei musculare în cașexia cancerului poate prelungi semnificativ viața (Zhou și colab., 2010). De asemenea, mulți indivizi mai în vârstă suferă de sarcopenie, o tulburare prelungită de pierdere a mușchilor care începe de obicei după vârsta de 50 de ani și implică o pierdere de aproximativ 1% din masa musculară pe an (Woo, 2017). Aceasta înseamnă că, până la vârsta de 80 de ani, indivizii sarcopenici au pierdut aproximativ 40% din masa musculară, un factor cheie în căderi, fragilitate și internări la domiciliu. În consecință, înțelegerea mecanismelor și a potențialelor răspunsuri terapeutice la atrofie este de interes general de bază și clinic (Ali și Garcia, 2014; Cohen și colab., 2015; Ziaaldini și colab., 2017).
ipoteza domeniului Mionuclear
mușchiul scheletic este destul de unic prin faptul că celulele mature sunt sincițiale și pot conține sute de nuclee. Aceasta este o adaptare necesară, având în vedere că fibrele musculare individuale pot fi enorme, atingând lungimi de până la ~600 mm (23 in) (de exemplu, mușchiul sartorius; Yang și colab., 1998) și volume care pot fi de peste 100.000 de ori mai mari decât o celulă mononucleată tipică (Bruusgaard și colab., 2003). Contribuțiile din mai multe nuclee sunt necesare pentru a produce cantități mari de ARNm necesare pentru a direcționa sinteza prodigioasă a proteinelor necesare pentru a genera și menține aparatul contractil (Nevalainen și colab., 2013).
atât natura plastică a mușchiului, cât și organizarea sa sincițială au dat naștere unei controverse care abia recent pare să fi fost rezolvată—”ipoteza domeniului mionuclear” (Qaisar și Larsson, 2014; Gundersen, 2016; Schwartz și colab., 2016). Această teorie își are originile în conceptul de „Wirkungssph Otrivre” sau „sferă de influență” propus de Strassburger (1893), în care susținea că un nucleu poate susține doar un volum discret de citoplasmă, definind astfel limitele superioare ale dimensiunii celulei. Acest principiu a fost elaborat în continuare de Gregory (2001) care a susținut că conținutul și volumul acidului dezoxiribonucleic celular sunt strâns cuplate. Natura sincițială a mușchilor scheletici ajută aceste celule să depășească această limitare. Ipoteza domeniului mionuclear dictează că, pentru a menține raportul nuclear-citoplasmatic adecvat, se adaugă noi nuclee în timpul hipertrofiei și se pierd cu atrofie.
există date substanțiale care demonstrează că numărul nuclear crește odată cu hipertrofia musculară (Moss, 1968; Cabric și James, 1983; Egner și colab., 2016), deși rămân unele controverse (Murach și colab., 2018). Acest proces a fost un domeniu de investigație intensă și unele dintre mecanismele moleculare subiacente care reglează achiziția de noi nuclee în timpul hipertrofiei au fost definite recent (Guerci și colab., 2012; Bentzinger și colab., 2014; Ross și colab., 2018). Acești nuclei supranumerari sunt dobândiți atunci când celulele stem restricționate prin descendență, de obicei celulele satelit, fuzionează cu fibrele musculare și contribuie cu nucleele lor (Brack și Rando, 2012; Bachman și colab., 2018). Celulele satelit se află sub membrana bazală și se învecinează cu sarcolema (Katz, 1961; Mauro, 1961). Acestea rămân în repaus până când sunt stimulate fie de steroizi anabolizanți, cum ar fi testosteronul, fie de leziuni focale în urma exercițiilor de rezistență, moment în care reintroduc ciclul celular și proliferează (Joubert și Tobin, 1995; Abreu și colab., 2017). Unele dintre celulele fiice fuzionează cu fibra musculară și facilitează atât repararea, cât și creșterea, în timp ce altele arestează și reconstituie piscina prin satelit (Dumont și colab., 2015; Goh și Millay, 2017). Celulele excedentare rămase suferă apoptoză (Schwartz, 2008).
aspectul controversat al ipotezei domeniului mionuclear este afirmația că mionucleii se pierd în timpul atrofiei. Există multe intervenții care induc atrofie la modelele animale, inclusiv: imobilizare, denervare și sepsis (Fitts și colab., 1986; Minnaard și colab., 2005; O ‘ Leary și colab., 2012). În fiecare caz, există o pierdere netă atât a zonei secțiunii transversale musculare (testul primar), cât și a apariției celulelor apoptotice în țesut (McCall și colab., 1998; Smith și colab., 2000; Strasser și colab., 2000; Alway și colab., 2003; McClung și colab., 2007; Andrianjafiniony și colab., 2010; Guo și colab., 2012; Palumbo și colab., 2012; Barnes și colab., 2015; Cheema și colab., 2015; Li și colab., 2016; Kletzien și colab., 2018). În aceste studii, apoptoza este măsurată într-o varietate de metode, inclusiv activarea caspazei, eliberarea endogului mitocondrial sau fragmentarea ADN-ului . Aceste studii oferă date convingătoare că apoptoza crește dramatic în timpul fazei timpurii a atrofiei. De exemplu, într-un studiu cuprinzător recent (Guo și colab., 2012), Guo și colab. șoarecii supuși la 14 zile de suspensie a membrelor posterioare, un tratament care a dus la o reducere de ~69% a greutății umede a mușchilor și la o reducere de ~43,8% a ariei secțiunii transversale în raport cu mușchiul contralateral nemanipulat. Autorii au cuantificat, de asemenea, un număr de markeri de apoptoză, inclusiv colorarea tunelului, scindarea/activarea caspazei-3 și scindarea poli(adenozin difosfat riboză) polimerază (PARP), o proteină implicată în repararea ADN-ului. Fiecare dintre aceste măsuri de apoptoză a crescut semnificativ după suspendarea membrelor posterioare. În timp ce accentul principal al acestei lucrări a fost de a evalua impactul pozitiv al stimulării electrice asupra limitării atrofiei în urma unei insulte, ei și mulți alți cercetători interpretează aceste tipuri de date ca oferind un sprijin puternic pentru ipoteza domeniului mionuclear.
dintr-o perspectivă biologică celulară, pierderea prezumtivă a nucleelor într-un țesut sincițial precum mușchiul scheletic prezintă o problemă practică majoră. Cum poate un nucleu individual să devină atât de compromis încât genomul său se condensează rapid și se fragmentează în timp ce vecinii săi persistă și ajută la menținerea viabilității fibrei musculare? Având în vedere că apoptoza este de obicei mediată de activarea clasei de proteaze cisteină cunoscute sub numele de caspaze, nu este clar ce mecanism ar putea servi la restricționarea activității unei protează difuzibile într-o citoplasmă comună.
această întrebare a fost abordată indirect într-un alt tip de celule sincițiale, sincitiotrofoblastul uman, un țesut care înconjoară placenta și conține aproximativ 5 nuclee 1010 (Mayhew și colab., 1999). Când apoptoza este indusă în sincitiotrofoblast, aceasta se propagă ca o undă la o rată de aproximativ 5 microni pe minut până când este implicat întregul țesut (Longtine și colab., 2012). În consecință, nu există regiuni „privilegiate” în citoplasma sincițială și toți nucleii sunt în cele din urmă distruși.
una dintre provocările cheie cu analiza apoptozei în mușchiul scheletic este că este un țesut foarte eterogen, unde aproximativ jumătate din nucleele sale se află în afara fibrelor musculare (Schmalbruch și Hellhammer, 1977). Aceste celule mononucleate includ celule satelit, celule endoteliale, fibroblaste, pericite și macrofage (Tedesco și colab., 2010). În consecință, este foarte dificil să se determine care parte a sarcolemului, se află un nucleu și, prin urmare, dacă este un adevărat mionucleus.
imagistica in Vivo Time-Lapse a fibrelor musculare marcate de șoarece
În ciuda numărului mare de lucrări care demonstrează apoptoza în timpul atrofiei musculare, mai mulți autori au pus la îndoială aceste rezultate (Wada și colab., 2002; Zhong și colab., 2005; Aravamudan și colab., 2006; Gundersen și Bruusgaard, 2008; Duddy și colab., 2011; Qaisar și Larsson, 2014). De exemplu, folosind fibre musculare izolate in vitro, Duddy și colab. a remarcat că, în timp ce volumul fibrelor musculare a scăzut în timp, reflectând un proces atrofic, numărul de mionuclei părea să fie stabil (Duddy și colab., 2011).
într-o serie de experimente elegante, Laboratorul Gundersen a injectat fibre musculare extensor digitorum longus (EDL) sau mușchii soleus cu coloranți care au etichetat independent nucleele și citoplasma la șoarecii anesteziați (Bruusgaard și Gundersen, 2008; Bruusgaard și colab., 2010). Acest lucru le-a permis să identifice fiecare nucleu în fibrele musculare individuale in vivo în timp și apoi să-și evalueze soarta. De exemplu, mușchii EDL au fost induși la hipertrofie prin ablația sinergilor lor majori (Bruusgaard și colab., 2010). Între zilele 6 și 11, Numărul de mionuclei a crescut cu aproximativ 54%, iar între zilele 9 și 14 a existat o creștere cu 35% a suprafeței secțiunii transversale (Figura 1). Aceste date sunt în concordanță cu ipoteza că mușchii dobândesc nuclee supranumerare înainte de creșterea majoră a fibrei în timpul hipertrofiei.

Figura 1. Myonucleii sunt dobândiți în timpul hipertrofiei, dar nu se pierd în timpul atrofiei la șoarece. Micrografele aceleiași fibre musculare EDL în timp după inducerea hipertrofiei (rândul de sus) și inducerea ulterioară a atrofiei (rândul de jos). Oligonucleotidele marcate fluorescent au fost utilizate pentru a vizualiza nucleele in vivo. Liniile punctate reprezintă sarcolema. Scara bar = 50 oktvm. (Adaptat de la Bruusgaard și colab., 2010. Folosit cu permisiunea procedurilor Academiei Naționale de științe.)
au examinat apoi soarta acestor mionuclei identificabile în mod unic în aceleași fibre musculare atunci când au fost induse să sufere atrofie. Denervarea a dus la o reducere de aproximativ 50% a volumului fibrelor musculare, dar nu a pierdut mionuclei (Figura 1; Bruusgaard și colab., 2010). Nerespectarea pierderii nucleare nu s-a datorat stimulului atrofic utilizat, deoarece au obținut aceleași rezultate atunci când mușchii au fost induși să atrofieze ca răspuns la blocarea nervului indusă de tetrodotoxină, suspensia membrelor posterioare, cașexia cancerului sau detraining (Bruusgaard și Gundersen, 2008; Bruusgaard și colab., 2010; Winje și colab., 2018a). De fapt, pe parcursul studiilor lor, au examinat mai mult de 200.000 de mionuclei individuali în mușchii atrofici și au observat doar 4 nuclee TUNEL-pozitive (apoptotice), ceea ce reprezintă o pierdere de doar ~0,002% din nuclee (Bruusgaard și colab., 2012). Aceste date susțin puternic ipoteza că atrofia musculară scheletică nu este însoțită de moartea mionucleară.
mușchiul insectelor ca Model
oricât de puternice sunt aceste studii, există unele limitări inerente utilizării modelelor de mamifere. În primul rând, se bazează invariabil pe intervenții experimentale, cum ar fi denervarea, pentru a induce atrofia musculară. În al doilea rând, mușchiul contralateral este de obicei folosit ca un control intern, chiar dacă și el este expus la unele dintre aceleași semnale de mediu, cum ar fi hormonii de stres (Bonaldo și Sandri, 2013). În cele din urmă, mușchii scheletici pot suferi, de asemenea, moarte celulară programată (PCD) în timpul dezvoltării, iar aceste modele abordează rareori rolul apoptozei în aceste celule.
un model alternativ care nu suferă de niciuna dintre aceste limitări și permite studiul nucleelor musculare atât în timpul atrofiei, cât și al PCD natural, este mușchiul intersegmental (ISM) din tutunul Hawkmoth Manduca sexta. ISM-urile sunt compuse din foi de fibre musculare uriașe, unde fiecare celulă are o lungime de aproximativ 5 mm și un diametru de până la 1 mm (figura 2a). ISM-urile se atașează la limitele segmentale din abdomen și generează atât comportamentul târâtor al larvelor, cât și comportamentul eclozion (emergență) al moliei adulte atunci când scapă din cuticula pupală la sfârșitul metamorfozei.
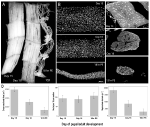
Figura 2. Retenția mionuclei atât în timpul atrofiei, cât și al morții mușchilor intersegmentali (ISMs) din molia Manduca sexta. (A) ISMs din trei etape de dezvoltare: homeostatic (zilele 15 de dezvoltare pupal-adult; stânga); atrofic (ziua 18; mijloc) ; și pe moarte (18 h post-ecloziune; dreapta). Bara de scară este egală cu ~1 mm (adaptat de la Schwartz și colab., 2016). (B) ISM-urile din aceleași etape de dezvoltare au fost eliminate și colorate cu colorantul nuclear DAPI și vizualizate prin microscopie confocală (adaptat de la Schwartz și colab., 2016). (C) secțiunile de fibre ISM (10 centimetri) au fost colorate cu colorantul nuclear DAPI. Rețineți pierderea dramatică a proteinelor musculare (zona gri deschis) în timpul atrofiei și morții, dar reținerea nucleelor în toate etapele (adaptat de la Schwartz și colab., 2016). (D) cuantificarea volumului fibrelor ISM (stânga), a numărului nuclear (mijloc) și a dimensiunii domeniului mionuclear (dreapta) în timpul homeostaziei, atrofiei și morții. (Eroare standard medie de la clasa a VIII-a.) (Adaptat de la Schwartz și colab., 2016).
În ziua 15 din cele 18 zile normale de dezvoltare pupal-adult, ISM-urile inițiază un program de atrofie declanșat hormonal care are ca rezultat o pierdere de masă de 40% până la ecloziune 3 zile mai târziu (figura 2a; Schwartz și Truman, 1983). Această pierdere dramatică a masei musculare este echivalentă cu cea observată la persoanele în vârstă de 80 de ani cu sarcopenie, dar are loc pe parcursul a 3 zile, mai degrabă decât 30 de ani. În timp ce ISM-urile pierd masă, ele păstrează proprietăți fiziologice normale, cum ar fi potențialul de repaus și forța/aria secțiunii transversale (Schwartz și Ruff, 2002). ISM-urile sunt folosite pentru a genera comportamentul eclozion la sfârșitul zilei 18 și apoi inițiază PCD, ambele fiind declanșate de un hormon peptidic (Schwartz și Truman, 1982, 1983). .spre deosebire de mușchii mamiferelor, ISM-urile nu conțin capilare, celule satelit, celule endoteliale sau pericite, ceea ce înseamnă că practic toate nucleele din țesut se află în fibrele musculare (Beaulaton și Lockshin, 1977). Am folosit două metode independente pentru a monitoriza soarta nucleară atât în timpul atrofiei, cât și al morții în acest model (Schwartz și colab., 2016). Prima a fost o abordare anatomică standard. Când ISM-urile suferă atrofie și moarte, există reduceri dramatice atât în volum, cât și în secțiunea transversală a fibrelor, dar în mod grosolan, numărul nuclear a rămas neschimbat (figura 2b). Folosind țesuturi secționate pentru Studii Cantitative, am constatat că a existat o reducere de ~49% a suprafeței secțiunii transversale a fibrelor în timpul atrofiei și o scădere suplimentară de 30% în primele etape ale morții (figurile 2C,D; Schwartz și colab., 2016). Cu toate acestea, numărul de nuclee nu s-a schimbat în aceeași perioadă, ceea ce înseamnă că a existat funcțional o reducere de 84% a domeniului mionuclear.
a doua metodă pe care am folosit-o a fost măsurarea conținutului de ADN al fibrelor musculare individuale de la animale în ziua 13, când mușchii sunt homeostatici, până la 18 ore post-ecloziune, când mușchii sunt foarte degenerați (Schwartz și colab., 2016). Am cuantificat conținutul de ADN al 420 de celule individuale și am constatat că nu a existat o pierdere semnificativă în timpul dezvoltării. Aceste analize biochimice sunt de acord cu datele anatomice și sugerează că concentrația relativă a nucleelor și a ADN-ului genomic a crescut funcțional pe măsură ce mușchii s-au atrofiat și au murit. În consecință, datele de la șoareci și molii sugerează că nucleele musculare nu suferă apoptoză în timpul atrofiei (sau PCD) și conduc la concluzia că ipoteza domeniului mionuclear ar trebui respinsă.
concluzii
aceste observații au o serie de implicații atât pentru înțelegerea biologiei de bază a mușchilor, cât și pentru dezvoltarea potențialelor intervenții terapeutice. Deși există date substanțiale care raportează prezența nucleelor apoptotice în țesut în urma insultelor atrofice, datele recente sugerează că acestea nu sunt adevărate mionuclei, ci mai degrabă celule mononucleare condamnate care se află în afara fibrei musculare. Motivul principal al acestei discrepanțe este că instrumentele tradiționale utilizate pentru detectarea apoptozei nu au rezoluția necesară pentru a determina în mod adecvat care parte a sarcolemului se află o celulă pe moarte. Metode care etichetează în mod specific myonuclei, fie prin injecții in vivo de coloranți fluorescenți (Bruusgaard și Gundersen, 2008), fie prin manipulări genetice pentru a exprima proteinele marker (Duddy și colab., 2011), dezvăluie puține nuclee apoptotice în fibrele etichetate. Demonstrația recentă că anticorpii direcționați împotriva proteinei Pericentriolare Material 1 (PCM1) etichetează numai myonuclei autentice oferă un instrument promițător care poate fi aplicat materialelor histologice tradiționale care sunt analizate în mod obișnuit în domeniu (Winje și colab., 2018b).
eșecul mușchilor scheletici de a folosi apoptoza ca mecanism de eliminare a mionuclei nu este surprinzător. Se pare că mușchiul scheletic matur este în mare măsură împiedicat să inițieze apoptoza prin faptul că reglează în sus expresia proteinelor puternice de supraviețuire, cum ar fi inhibitorul legat de X al proteinei de apoptoză (Xiap) (Smith și colab., 2009) și apoptoza represor cu domeniul de recrutare caspase (ARC) (Xiao și colab., 2011), exprimând simultan niveluri scăzute de mediatori de apoptoză, cum ar fi factorul 1 de activare a proteazei apoptotice (Apaf-1) (Burgess și colab., 1999). Acest lucru are sens teleologic, deoarece mușchii sunt supuși unor perturbații extreme, inclusiv perturbarea sarcolemului după exerciții intense, deci ar fi catastrofal dacă ar declanșa moartea celulară, mai degrabă decât să inițieze repararea țesuturilor și, eventual, hipertrofia. Ca și alte celule diferențiate terminal, mușchii tind să folosească mecanisme non-apoptotice, mai ales PCD autofagic (cunoscut și sub numele de degenerare de tip II) (Clarke, 1990; Schwartz și colab., 1993; Kole și colab., 2013; Ginet și colab., 2014) când mor în timpul dezvoltării.
chiar dacă ar exista un mecanism prin care mușchii ar putea viza selectiv nucleele individuale, nu este clar că acest lucru ar fi benefic. De fapt, reținerea nucleelor „excedentare” în timpul atrofiei conferă un avantaj distinct individului, deoarece mușchii scheletici suferă frecvent cicluri de atrofie și hipertrofie ca răspuns la condițiile de mediu, cum ar fi disponibilitatea alimentelor. Capacitatea de a recupera rapid prin utilizarea myonuclei preexistente poate servi un rol important în adaptare (Jackson și colab., 2012) și ajută la explicarea fenomenului „memoriei musculare” (Staron și colab., 1991; Gundersen, 2016). Este bine documentat în domeniul fiziologiei exercițiilor că este mult mai ușor să redobândești un anumit nivel de fitness muscular prin exerciții fizice decât a fost să-l atingi pe primul loc, chiar dacă a existat o perioadă lungă de intervenție de detraining. Cu alte cuvinte, sintagma „folosește-l sau pierde-l” poate fi articulată mai precis ca „folosește-l sau pierde-l, până când lucrezi din nou la el.”Acest lucru a fost demonstrat direct de un alt experiment de la laboratorul Gundersen care a demonstrat că, odată ce un mușchi a dobândit nuclee noi, le reține mult timp după îndepărtarea stimulului hipertrofic. Au indus hipertrofia musculară la șoarecii femele tratându-i timp de 2 săptămâni cu testosteron și apoi au examinat mușchii la 3 săptămâni după retragerea steroizilor (Egner și colab., 2013). Volumul muscular a revenit la valoarea inițială, dar nucleele nou dobândite au persistat chiar și 3 luni mai târziu. Când mușchii au fost supuși supraîncărcării pentru a reinițializa hipertrofia, cei tratați cu steroizi au suferit rapid o creștere cu 36% a volumului de fibre, în timp ce mușchii de control au crescut doar cu 6%. Aceste date sugerează că nucleele” excedentare ” ar putea fi mobilizate rapid pentru a facilita recalificarea.
aceste observații au implicații potențiale pentru politica de sănătate publică. S-a demonstrat că creșterea musculară, funcția fiziologică și capacitatea regenerativă scad odată cu vârsta, în mare parte datorită proliferării reduse a celulelor satelit (Blau și colab., 2015). În consecință, exercițiile fizice în timpul adolescenței, când creșterea musculară este sporită de hormoni, nutriție și o piscină robustă prin satelit, ar putea servi funcțional pentru a permite persoanelor să „banculeze” mionuclei care ar putea fi atrase mai târziu în viață pentru a încetini efectele îmbătrânirii și, eventual, pentru a preveni sarcopenia.
în plus, aceste date au implicații în domeniul sporturilor competitive. Utilizarea steroizilor anabolizanți este un stimul puternic pentru hipertrofia musculară și adăugarea de noi mionuclei (Egner și colab., 2013). Deoarece aceste nuclee persistă mult timp după terminarea utilizării steroizilor, sportivii obțin probabil beneficiile consumului ilegal de droguri fără riscul de detectare.în rezumat, în timp ce adăugarea de noi nuclee cu creștere musculară este în mare măsură acceptată, pierderea apoptotică a nucleelor cu atrofie nu poate fi susținută, sugerând că interpretarea strictă a ipotezei domeniului mionuclear nu poate fi susținută. În schimb, se pare că, odată dobândite, myonucleii persistă chiar și atunci când un mușchi devine atrofic sau inițiază moartea celulelor.
contribuții autor
autorul confirmă că este singurul contribuitor al acestei lucrări și a aprobat-o pentru publicare.
finanțare
lucrarea pentru această publicație a fost susținută de fonduri de la Eugene M. și Ronnie Isenberg Professorship Endowment. Nici un sprijin corporativ a fost oferit autorului și nici un produs comercial sunt menționate sau aprobate.
Declarație privind conflictul de interese
autorul declară că cercetarea a fost realizată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
mulțumiri
autorul ar dori să-i mulțumesc doamnei Christine Brown pentru asistență în pregătirea acestui manuscris și Eugene M. și Ronnie Isenberg Professorship Endowment pentru sprijin.
Aagaard, P. (2004). Efectuarea mușchilor „mai puternici”: exerciții fizice, nutriție, medicamente. J. Musculoskelet. Neuronale. Interacționează. 4, 165–174.
PubMed Abstract/Google Scholar
Abreu, P., Mendes, S. V., Ceccatto, V. M. și Hirabara, S. M. (2017). Activarea celulelor prin satelit indusă de adaptarea musculară aerobă ca răspuns la exercițiile de anduranță la oameni și rozătoare. Life Sci. 170, 33–40. doi:10.1016 / j. if.2016.11.016
PubMed Abstract | CrossRef Full Text/Google Scholar
Ali, S. și Garcia, J. M. (2014). Sarcopenie, cașexie și îmbătrânire: diagnostic, mecanisme și opțiuni terapeutice—o mini-revizuire. Gerontologie 60, 294-305. doi: 10.1159 / 000356760
PubMed Abstract | CrossRef Full Text/Google Scholar
Alway, S. E., Martyn, J. K., Ouyang, J., Chaudhrai, A. și Murlasits, Z. S. (2003). Id2 expression during apoptosis and satellite cell activation in unloaded and loaded quail skeletal muscles. Am. J. Physiol. Regul. Integr. Comp. Physiol. 284, R540–R549. doi: 10.1152/ajpregu.00550.2002
PubMed Abstract | CrossRef Full Text | Google Scholar
Andrianjafiniony, T., Dupre-Aucouturier, S., Letexier, D., Couchoux, H., and Desplanches, D. (2010). Oxidative stress, apoptosis, and proteolysis in skeletal muscle repair after unloading. Am. J. Phys. Cell Physiol. 299, C307–C315. doi: 10.1152/ajpcell.00069.2010
PubMed Abstract | CrossRef Full Text/Google Scholar
Aravamudan, B., Mantilla, C. B., Zhan, W. Z. și Sieck, G. C. (2006). Efecte de denervare asupra dimensiunii domeniului mionuclear al fibrelor diafragmei de șobolan. J. Appl. Physiol. 100, 1617–1622. doi: 10.1152 / japplphysiol.01277.2005
CrossRef Full Text/Google Scholar
Bachman, J. F., Klose, A., Liu, W., Paris, N. D., Blanc, R. S., Schmalz, M. și colab. (2018). Creșterea prepubertală a mușchilor scheletici necesită contribuția mionucleară derivată din celulele satelit care exprimă Pax 7. Dezvoltare 145: 167197.
Google Scholar
Barnes, B. T., Confides, A. L., Rich, M. M. și Dupont-Versteegden, E. E. (2015). Căile apoptotice musculare distincte sunt activate în mușchii cu diferite tipuri de fibre într-un model de șobolan de miopatie critică a bolii. J. Mușchi Res. Cell Motil. 36, 243–253. doi: 10.1007 / s10974-015-9410-8
PubMed Abstract | CrossRef Full Text/Google Scholar
Beaulaton, J. și Lockshin, R. A. (1977). Studiul Ultrastructural al degenerării normale a mușchilor intersegmentali ai Anthereae polyphemus și Manduca sexta (insecta, lepidoptera) cu referire specială la autofagia celulară. J. Morphol. 154, 39–57. doi: 10.1002 / jmor.1051540104
PubMed Abstract | CrossRef Full Text/Google Scholar
Bentzinger, C. F., von Maltzahn, J., Dumont, N. A., Stark, D. A., Wang, Y. X., Nhan, K. și colab. (2014). Wnt 7a stimulează motilitatea și grefarea celulelor stem miogene, rezultând o forță musculară îmbunătățită. J. Cell Biol. 205, 97–111. doi: 10.1083 / jcb.201310035
PubMed Abstract | CrossRef Full Text/Google Scholar
Blau, H. M., Cosgrove, B. D. și Ho, A. T. (2015). Rolul central al celulelor stem musculare în eșecul regenerativ cu îmbătrânirea. Nat. Med. 21, 854–862. doi: 10.1038 / nm.3918
PubMed Abstract | CrossRef Full Text/Google Scholar
Bonaldo, P. și Sandri, M. (2013). Mecanisme celulare și moleculare ale atrofiei musculare. Dis. Model. Mech. 6, 25–39. doi: 10.1242 / dmm.010389
CrossRef Full Text/Google Scholar
Brack, A. S. și Rando, T. A. (2012). Celule stem specifice țesutului: lecții din celula satelit a mușchilor scheletici. Celula Stem 10, 504-514. doi: 10.1016/j.tulpină.2012.04.001
PubMed Abstract | CrossRef Full Text/Google Scholar
Bruusgaard, J. C., Egner, I. M., Larsen, T. K., Dupre-Aucouturier, S., Desplanches, D., și Gundersen, K. (2012). Nici o modificare a numărului mionuclear în timpul descărcării și reîncărcării musculare. J. Appl. Physiol. 113, 290–296. doi: 10.1152 / japplphysiol.00436.2012
PubMed Abstract | CrossRef Full Text/Google Scholar
Bruusgaard, J. C., și Gundersen, K. (2008). Microscopia in vivo time-lapse nu relevă nicio pierdere de mionuclei murine în timpul săptămânilor de atrofie musculară. J. Clin. Investește. 118, 1450–1457. doi: 10.1172 / JCI34022
PubMed Abstract | CrossRef Full Text/Google Scholar
Bruusgaard, J. C., Johansen, I. B., Egner, I. M., Rana, Z. A. și Gundersen, K. (2010). Myonucleii dobândiți prin exerciții de suprasarcină preced hipertrofia și nu e pierd pe detraining. Proc. Natl. Acad. Sci. SUA 107, 15111-15116. doi: 10.1073 / pnas.0913935107
CrossRef Full Text/Google Scholar
Bruusgaard, J. C., Liestol, K., Ekmark, M., Kollstad, K. și Gundersen, K. (2003). Numărul și distribuția spațială a nucleelor în fibrele musculare ale șoarecilor normali studiați in vivo. J. Physiol. 551, 467–478. doi: 10.1113 / jphysiol.2003.045328
PubMed Abstract | CrossRef Full Text/Google Scholar
Burgess, D. H., Svensson, M., Dandrea, T., Gronlund, K., Hammarquist, F., Orrenius, S. și colab. (1999). Citosolii musculaturii scheletice umane sunt refractari la activarea dependentă de citocrom C a caspazelor de tip II și nu au APAF-1. Moartea Celulelor Diferă. 6, 256–261. doi: 10.1038 / sj.cdd.4400489
PubMed Abstract | CrossRef Full Text/Google Scholar
Cabric, M. și James, N. T. (1983). Analize morfometrice asupra mușchilor câinilor antrenați și neinstruiți. Am. J. Anat. 166, 359–368. doi: 10.1002 / aja.1001660309
PubMed Abstract | CrossRef Full Text/Google Scholar
Cheema, N., Herbst, A., McKenzie, D. și Aiken, J. M. (2015). Apoptoza și necroza mediază pierderea fibrelor musculare scheletice în anomaliile enzimatice mitocondriale induse de vârstă. Celula De Îmbătrânire 14, 1085-1093. doi: 10.1111 / acela.12399
PubMed Abstract | CrossRef Full Text/Google Scholar
Clarke, P. G. (1990). Moartea celulelor de dezvoltare: diversitate morfologică și mecanisme multiple. Anat. Embryol. 181, 195–213.
Google Scholar
Cohen, S., Nathan, J. A. și Goldberg, A. L. (2015). Pierderea musculară în boală: mecanisme moleculare și terapii promițătoare. Nat. Rev. Droguri Discov. 14, 58–74. doi: 10.1038 / nrd4467
PubMed Abstract | CrossRef Full Text/Google Scholar
Duddy, W. J., Cohen, T., Duguez, S. și Partridge, T. A. (2011). Fibra musculară izolată ca model de atrofie neutilizată: caracterizare folosind PhAct, o metodă de cuantificare a f-actinei. Exp. Rezoluția Celulei 317, 1979-1993. doi: 10.1016 / j. yexcr.2011.05.013
PubMed Abstract | CrossRef Full Text/Google Scholar
Dumont, N. A., Wang, Y. X. și Rudnicki, ma (2015). Mecanisme intrinseci și extrinseci care reglează funcția celulelor satelit. Dezvoltare 142, 1572-1581. doi: 10.1242 / dev.114223
PubMed Abstract | CrossRef Full Text/Google Scholar
Egner, I. M., Bruusgaard, J. C., Eftestol, E., și Gundersen, K. (2013). Un mecanism de memorie celulară ajută la hipertrofia supraîncărcării în mușchi mult timp după o expunere episodică la steroizi anabolizanți. J. Physiol. 591, 6221–6230. doi: 10.1113 / jphysiol.2013.264457
PubMed Abstract | CrossRef Full Text/Google Scholar
Egner, I. M., Bruusgaard, J. C. și Gundersen, K. (2016). Epuizarea celulelor prin satelit previne hipertrofia fibrelor în mușchii scheletici. Dezvoltare 143, 2898-2906. doi: 10.1242 / dev.134411
PubMed Abstract / CrossRef Full Text / Google Scholar
Fitts, R. H., Metzger, J. M., Riley, D. A. și Unsworth, B. R. (1986). Modele de neutilizare: o comparație a suspensiei posterioare și a imobilizării. J. Appl. Physiol. 60, 1946–1953.
Google Scholar
Ginet, V., Pittet, M. P., Rummel, C., Osterheld, M. C., Meuli, R., Clarke, P. G. și colab. (2014). Neuronii morți în talamusul nou-născuților și șobolanilor asfixiați sunt autofagi. Ann. Neurol. 76, 695–711. doi: 10.1002 / ana.24257
PubMed Abstract | CrossRef Full Text/Google Scholar
Goh, Q. și Millay, D. P. (2017). Cerința fuziunii celulelor stem mediate de miomaker pentru hipertrofia mușchilor scheletici. elife 6. doi: 10.7554 / eLife.20007
PubMed Abstract | CrossRef Full Text/Google Scholar
Gregory, T. R. (2001). Coincidență, coevoluție sau cauzalitate? Conținutul ADN – ului, dimensiunea celulei și enigma valorii C. Biol. Rev. Camb. Filos. Soc. 76, 65–101. doi: 10.1017/s1464793100005595
PubMed Abstract | CrossRef Full Text/Google Scholar
Guerci, A., Lahoute, C., Hebrard, S., Collard, L., Graindorge, D., Favier, M., și colab. (2012). Semnalele paracrine dependente de Srf produse de miofibre controlează hipertrofia musculară scheletică mediată de celule prin satelit. Metab Celulă. 15, 25–37. doi: 10.1016 / j.cmet.2011.12.001
PubMed Abstract | CrossRef Full Text/Google Scholar
Gundersen, K. (2016). Memorie musculară și un nou model celular pentru atrofie musculară și hipertrofie. J. Exp. Biol. 219, 235–242. doi: 10.1242/jeb.124495
PubMed Abstract | CrossRef Full Text/Google Scholar
Gundersen, K. și Bruusgaard, J. C. (2008). Domenii nucleare în timpul atrofiei musculare: nucleele pierdute sau paradigma pierdută? J. Physiol. 586, 2675–2681. doi: 10.1113 / jphysiol.2008.154369
PubMed Abstract | CrossRef Full Text/Google Scholar
Guo, B. S., Cheung, K. K., Yeung, SS, Zhang, B. T. și Yeung, E. W. (2012). Stimularea electrică influențează proliferarea celulelor prin satelit și apoptoza în atrofia musculară indusă de descărcare la șoareci. PLoS Unul 7: e30348. doi: 10.1371/jurnal.pone.0053523
PubMed Abstract | CrossRef Full Text/Google Scholar
Jackson, J. R., Mula, J., Kirby, T. J., Fry, C. S., Lee, J. D., Ubele, M. F. și colab. (2012). Depleția celulelor prin satelit nu inhibă regenerarea mușchilor scheletici adulți după atrofia indusă de descărcare. Am. J. Phys. Fiziol Celular. 303, C854-C861. doi: 10.1152 / ajpcell.00207.2012
PubMed Abstract | CrossRef Full Text/Google Scholar
Joubert, Y. și Tobin, C. (1995). Tratamentul cu testosteron are ca rezultat activarea celulelor satelit în repaus și recrutarea în ciclul celular în mușchiul levator ani de șobolan. Dev. Biol. 169, 286–294. doi: 10.1006 / dbio.1995.1144
PubMed Abstract | CrossRef Full Text/Google Scholar
Katz, B. (1961). Terminațiile fibrei nervoase aferente în fusul muscular al broaștei. Filos. Trans. R. Soc. Lond. Biol. Sci. 243, 221–240.
PubMed Abstract/Google Scholar
Kletzien, H., Hare, aj, Leverson, G. și Connor, N. P. (2018). Efectul legat de vârstă al morții celulare asupra morfologiei și numărului fibrelor în mușchiul limbii. Nervul Muscular 57, E29-E37. doi: 10.1002 / mus.25671
PubMed Abstract | CrossRef Full Text/Google Scholar
Kole, A. J., Annis, R. P. și Deshmukh, M. (2013). Neuroni maturi: echipați pentru supraviețuire. Moartea Celulelor Dis. 4: e689. doi: 10.1038 / cddi.2013.220
PubMed Abstract | CrossRef Full Text/Google Scholar
Li, T. S., Shi, H., Wang, L. și Yan, C. (2016). Efectul celulelor stem mezenchimale ale măduvei osoase asupra proliferării celulelor satelit și a apoptozei în atrofia musculară indusă de imobilizare la șobolani. Med. Sci. Monit. 22, 4651–4660. doi: 10.12659/MSM.898137
PubMed Abstract | CrossRef Full Text/Google Scholar
Lockshin, R. A. și Williams, C. M. (1965). Moartea celulară programată – I. citologia degenerării în mușchii intersegmentali ai Mătăsii Pernyi. J. Physiol De Insecte. 11, 123–133. doi: 10.1016/0022-1910(65)90099-5
PubMed Abstract | CrossRef Full Text | Google Scholar
Longtine, ms, Barton, A., Chen, B. și Nelson, D. M. (2012). Imagistica cu celule vii arată că apoptoza inițiază local și se propagă ca un val pe tot parcursul sincitiotrofoblastelor în culturile primare ale trofoblastelor viloase placentare umane. Placenta 33, 971-976. doi: 10.1016 / j.placenta.2012.09.013
PubMed Abstract | CrossRef Full Text/Google Scholar
Mauro, A. (1961). Celula satelit a fibrelor musculare scheletice. J. Biophys. Biochem. Cytol. 9, 493–495.
Google Scholar
Mayhew, T. M., Leach, L., McGee, R., Ismail, W. W., Myklebust, R. și Lammiman, M. J. (1999). Proliferarea, diferențierea și apoptoza în trofoblastul vilos la 13-41 săptămâni de gestație (inclusiv observații privind lamelele anulate și complexele porilor nucleari). Placenta 20, 407-422. doi: 10.1053 / loc.1999.0399.
PubMed Abstract | CrossRef Full Text/Google Scholar
McCall, G. E., Allen, D. L., Linderman, J. K., Grindeland, R. E., Roy, R. R., Mukku, V. R., și colab. (1998). Menținerea dimensiunii domeniului mionuclear la șobolanul soleus după suprasarcină și tratamentul cu hormon de creștere/IGF-I. J. Appl. Physiol. 84, 1407–1412.
Google Scholar
McClung, J. M., Kavazis, A. N., DeRuisseau, K. C., Falk, D. J., Deering, ma, Lee, Y. și colab. (2007). Reglarea caspazei-3 a domeniului mionuclear al diafragmei în timpul atrofiei induse de ventilație mecanică. Am. J. Respir. Crit. Care Med. 175, 150–159. doi: 10.1164 / rccm.200601 – 142oc
PubMed Abstract | CrossRef Full Text/Google Scholar
Metter, E. J., Talbot, L. A., Schrager, M., și Conwit, R. (2002). Forța musculară scheletică ca predictor al mortalității din toate cauzele la bărbații sănătoși. J. Gerontol. Un Biol. Sci. Med. Sci. 57, B359-B365. doi: 10.1093 / gerona / 57.10.B359
PubMed Abstract | CrossRef Full Text/Google Scholar
Minnaard, R., Drost, Mr, Wagenmakers, A. J., van Kranenburg, G. P., Kuipers, H. și Hesselink, M. K. (2005). Pierderea mușchilor scheletici și performanța contractilă la șobolanii septici. Nervul Muscular 31, 339-348. doi: 10.1002 / mus.20268
PubMed Abstract | CrossRef Full Text/Google Scholar
Moss, F. P. (1968). Relația dintre dimensiunile fibrelor și numărul de nuclee în timpul creșterii normale a mușchilor scheletici la păsările domestice. Am. J. Anat. 122, 555–563. doi: 10.1002 / aja.1001220308
PubMed Abstract | CrossRef Full Text/Google Scholar
Murach, K. A., Englund, D. A., Dupont-Versteegden, E. E., McCarthy, J. J. și Peterson, C. A. (2018). Flexibilitatea domeniului mionuclear provoacă ipoteze rigide privind contribuția celulelor satelit la hipertrofia fibrelor musculare scheletice. În față. Physiol. 9:635. doi: 10.3389 / fphys.2018.00635
CrossRef Full Text/Google Scholar
Nevalainen, M., Kaakinen, M. și Metsikko, K. (2013). Distribuția transcrierilor ARNm și a activității de traducere în miofibre scheletice. Țesutul Celular Rez. 353, 539-548. doi: 10.1007 / s00441-013-1659-x
PubMed Abstract | CrossRef Full Text/Google Scholar
O ‘ Leary, M. F., Vainshtein, A., Carter, H. N., Zhang, Y. și Hood, D. A. (2012). Disfuncția mitocondrială indusă de denervare și autofagia în mușchiul scheletic al animalelor cu deficit de apoptoză. Am. J. Phys. Fiziol Celular. 303, C447–C454. doi: 10.1152/ajpcell.00451.2011
PubMed Abstract | CrossRef Full Text | Google Scholar
Palumbo, C., Rovesta, C., and Ferretti, M. (2012). Striated muscle fiber apoptosis after experimental tendon lesion in a rat model. J. Anat. 221, 358–363. doi: 10.1111/j.1469-7580.2012.01554.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Qaisar, R., and Larsson, L. (2014). What determines myonuclear domain size? Indian J. Physiol. Pharmacol. 58, 1–12.
PubMed Abstract | Google Scholar
Ross, J. A., Levy, Y., Svensson, K., Philp, A., Schenk, S. și Ochala, J. (2018). SIRT1 reglează numărul nuclear și dimensiunea domeniului în fibrele musculare scheletice. J. Cell. Physiol. 233, 7157–7163. doi: 10.1002 / jcp.26542
PubMed Abstract | CrossRef Full Text/Google Scholar
Schiaffino, S., Dyar, K. A., Ciciliot, S., Blaauw, B. și Sandri, M. (2013). Mecanisme care reglează creșterea și atrofia mușchilor scheletici. Februarie J. 280, 4294-4314. doi: 10.1111/februarie.12253
PubMed Abstract | CrossRef Full Text/Google Scholar
Schmalbruch, H. și Hellhammer, U. (1977). Numărul de nuclee din mușchii șobolanilor adulți, cu referire specială la celulele satelit. Anat. Rec. 189, 169–175. doi: 10.1002/ar.1091890204
PubMed Abstract | CrossRef Full Text/Google Scholar
Schwartz, L. M. (2008). Atrofia și moartea celulară programată a mușchilor scheletici. Moartea Celulelor Diferă. 15, 1163–1169. doi: 10.1038 / cdd.2008.68
PubMed Abstract | CrossRef Full Text/Google Scholar
Schwartz, L. M., Brown, C., McLaughlin, K., Smith, W. și Bigelow, C. (2016). Domeniul mionuclear nu este menținut în mușchiul scheletic nici în timpul atrofiei, nici în timpul morții celulare programate. Am. J. Phys. Fiziol Celular. 311, C607–C615. doi: 10.1152 / ajpcell.00176.2016
PubMed Abstract | CrossRef Full Text/Google Scholar
Schwartz, L. M. și Ruff, R. L. (2002). Modificări ale proprietăților contractile ale mușchilor scheletici în timpul atrofiei programate în dezvoltare și a morții. Am. J. Phys. Fiziol Celular. 282, C1270-C1277. doi: 10.1152 / ajpcell.01275.2000
CrossRef Full Text/Google Scholar
Schwartz, L. M., Smith, S. W., Jones, M. E., și Osborne, B. A. (1993). Toate decesele celulare programate apar prin apoptoză? Proc. Natl. Acad. Sci. SUA 90, 980-984.
Google Scholar
Schwartz, L. M. și Truman, J. W. (1983). Controlul Hormonal al ratelor de dezvoltare metamorfică în viermele de tutun Manduca sexta. Dev. Biol. 99, 103–114. doi: 10.1016/0012-1606(83)90257-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Schwartz, L. M. și Truman, J. W. (1982). Reglarea peptidică și steroidă a degenerării musculare la o insectă. Știință 215, 1420-1421. doi: 10.1126 / știință.6278594
PubMed Abstract | CrossRef Full Text/Google Scholar
Smith, H. K., Maxwell, L., Martyn, J. A. și Bass, J. J. (2000). Fragmentarea ADN-ului Nuclear și modificările morfologice ale mușchilor scheletici ai iepurilor adulți după imobilizarea pe termen scurt. Țesut Celular Res. 302, 235-241. doi: 10.1007/s004410000280
PubMed Abstract | CrossRef Full Text/Google Scholar
Smith, M. I., Huang, Y. Y. și Deshmukh, M. (2009). Diferențierea mușchilor scheletici evocă XIAP endogen pentru a restricționa calea apoptotică. PLoS Unul 4: e5097. doi: 10.1371 / jurnal.pone.0005097
CrossRef Full Text/Google Scholar
Staron, R. S., Leonardi, M. J., Karapondo, D. L., Malicky, E. S., Falkel, J. E., Hagerman, F. C. și colab. (1991). Rezistență și adaptări ale mușchilor scheletici la femeile instruite cu rezistență grea după detraining și recalificare. J. Appl. Physiol. 70, 631-640.
Google Scholar
Strassburger, E. (1893). Sfera de acțiune a nucleelor și dimensiunea celulei. Histol Beitr. 5, 97-124.
Google Scholar
Strasser, H., Tiefenthaler, M., Steinlechner, M., Eder, I., Bartsch, G. și Konwalinka, G. (2000). Apoptoza dependentă de vârstă și pierderea celulelor rhabdosphincter. J. Urol. 164, 1781–1785. doi: 10.1016/S0022-5347(05)67106-6
PubMed Abstract | CrossRef Full Text/Google Scholar
Tedesco, F. S., Dellavalle, A., Diaz-Manera, J., Messina, G. și Cossu, G. (2010). Repararea mușchilor scheletici: potențialul regenerativ al celulelor stem ale mușchilor scheletici. J. Clin. Investește. 120, 11–19. doi: 10.1172 / JCI40373
PubMed Abstract | CrossRef Full Text/Google Scholar
Wada, K. I., Takahashi, H., Katsuta, S., și soia, H. (2002). Nu există o scădere a numărului mionuclear după denervarea pe termen lung la șoarecii maturi. Am. J. Phys. Fiziol Celular. 283, C484–C488. doi: 10.1152 / ajpcell.00025.2002
CrossRef Full Text/Google Scholar
Winje, I. M., Sheng, X., Hansson, K. A., Solbr, A., Tenn, S., Saatcioglu, F. și colab. (2018a). Cașexia nu induce pierderea de mionuclei sau fibre musculare în timpul cancerului de prostată xenogrefat la șoareci. Acta Physiol. e13204. doi: 10.1111 / apha.13204
CrossRef Full Text/Google Scholar
Winje, I. M., Bengtsen, M., Eftestol, E., Juvkam, I., Bruusgaard, J. C. și Gundersen, K. (2018b). Etichetarea specifică a mionuclei de către un anticorp împotriva materialului pericentriolar 1 pe secțiunile țesutului muscular scheletic. Acta Physiol. 223: e13034. doi: 10.1111 / apha.13034
CrossRef Text Complet/Google Scholar
Woo, J. (2017). Sarcopenie. Clin. Geriatr. Med. 33, 305–314. doi: 10.1016 / j. cger.2010.02.012
CrossRef Full Text/Google Scholar
Xiao, R., Ferry, A. L. și Dupont-Versteegden, E. E. (2011). Rezistența la moarte celulară a miotuburilor diferențiate este asociată cu mecanisme anti-apoptotice îmbunătățite în comparație cu mioblastele. Apoptoza 16, 221-234. doi: 10.1007 / s10495-010-0566-9
PubMed Abstract | CrossRef Full Text/Google Scholar
Yang, D., Morris, S. F. și Sigurdson, L. (1998). Mușchiul sartorius: considerații anatomice pentru chirurgii reconstructivi. Chirurg Radiol. Anat. 20, 307–310. doi: 10.1007
PubMed Abstract | CrossRef Full Text/Google Scholar
Zhong, H., Roy, R. R., Siengthai, B. și Edgerton, V. R. (2005). Efectele inactivității asupra dimensiunii fibrelor și a numărului mionuclear în mușchiul soleus de șobolan. J. Appl. Physiol. 99, 1494–1499. doi: 10.1152 / japplphysiol.00394.2005
CrossRef Full Text/Google Scholar
Zhou, X., Wang, J. L., Lu, J., Song, Y., Kwak, K. S., Jiao, Q., și colab. (2010). Inversarea cașexiei cancerului și a pierderii musculare prin antagonismul act RIIB duce la supraviețuire prelungită. Celula 142, 531-543. doi: 10.1016 / j.celulă.2010.07.011
PubMed Abstract | CrossRef Full Text/Google Scholar
Ziaaldini, M. M., Marzetti, E., Picca, A. și Murlasits, Z. (2017). Căile biochimice ale sarcopeniei și modularea lor prin exerciții fizice: o revizuire narativă. În față. Med. 4:167. doi: 10.3389 / fmed.2017.00167
CrossRef Text Complet / Google Scholar