introducere
politica internațională de conservare recunoaște biodiversitatea la trei niveluri, ecosistem, specii și genetice și că managementul ar trebui să urmărească păstrarea tuturor celor trei (Convenția privind diversitatea biologică, 2007). Acest lucru se reflectă în mod clar în Convenția privind diversitatea biologică, Aichi Biodiversity Targets, convenită în 2010 (https://www.cbd.int/sp/targets/), unde există o referință specifică în obiective și ținte, nu numai la ecosisteme și specii, ci și la diversitatea genetică. Cu toate acestea, abordările actuale ale conservării biodiversității se bazează în mare parte pe zone geografice, ecosisteme, comunități ecologice și specii, cu mai puțină atenție asupra diversității genetice și a continuumului specie-populație. Într-adevăr, majoritatea practicienilor de conservare se concentrează pe specii ca metrică a biodiversității, luând în considerare unele unități intraspecifice din cadrul speciilor amenințate gestionate intens. În acest context, există o dorință naturală de a folosi Categorii simple—specii numite și alți taxoni numiți (adică subspecii) și, uneori, unități de conservare în cadrul speciilor. Speciile oferă, în special, o măsură comună a biodiversității care stă la baza multor eforturi științifice și de gestionare (Mace, 2004; Wilson, 2017). Ca unitate fundamentală de conservare, speciile sunt adesea folosite pentru a cuantifica valoarea biodiversității prin liste de specii din ariile protejate, identificarea speciilor amenințate în jurisdicții și ca bază pentru procedurile de biosecuritate. În plus, speciile sunt de obicei entitățile cu care comunicăm publicului problemele de conservare.
acest impuls de a identifica pur și simplu unități discrete în natură contravine mai multor realități înrudite. În primul rând, speciația nu este instantanee. Mai degrabă este de obicei un proces prelungit (Dynesius și Jansson, 2014; Roux și colab., 2016), progresând de la populații izolate, probabil supuse unor presiuni de selecție diferite, la specii lipsite de ambiguitate pe scări lungi de timp, cu inversări frecvente cauzate de schimbările de mediu sau de simpla fuziune genetică pe măsură ce intervalele se reconectează (Figura 1; Seehausen și colab., 2008; Rosenblum și colab., 2012). În al doilea rând, majoritatea speciilor recunoscute nu sunt uniforme genetic. Divergența (derivă și izolare prelungită) și reticularea (geneflow, hibridizare, introgresie și recombinare) modelează în mod eficient variația geografică și genomică în cadrul speciilor (Cutter și Payseur, 2013; Edwards și colab., 2016) și unele specii pot fi structurate în populații izolate lungi, dintre care unele ar putea justifica recunoașterea ca specii complete, dar poate criptice morfologic (Bickford și colab., 2007; Jorger și Schr, 2013). În al treilea rând, atât timp cât sunt recunoscute, speciile numite nu sunt echivalente în ceea ce privește câtă moștenire evolutivă reprezintă—pur și simplu, un ornitorinc (ca reprezentant al unei descendențe divergente antice) poate fi considerat a valora mai mult decât un rozător divergent recent (Isaac și colab., 2007). În al patrulea rând, denumirile speciilor nu sunt aruncate în piatră, ci mai degrabă sunt ipoteze care trebuie testate pe măsură ce devin disponibile noi dovezi (Hei și colab., 2003). Fiecare dintre aceste probleme creează complexitate și ambiguitate în delimitarea speciilor în general și atribuirea atenției de conservare acestora sau populațiilor lor componente. În plus, delimitarea speciilor este, de asemenea, supusă diferitelor concepte de specii și modalități de diagnosticare a acestora, ceea ce a provocat o mare consternare în rândul biologilor de conservare (Mace, 2004; Frankham și colab., 2012; Garnett și Christidis, 2017).
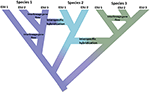
Figura 1. Speciația ca un continuum care progresează de la populații izolate la specii lipsite de ambiguitate, demonstrat de schimbarea culorii cu culorile reprezentând diferite specii. Unele linii care apar din cauza izolării populației se dezvoltă în specii adevărate, nodurile interne ale arborelui reprezentând inițierea procesului de speciație. Alte linii nu se dezvoltă în specii adevărate, dar diferă în timp, indicând faptul că speciile nu sunt uniforme genetic. Aceste linii sunt prezentate ca unități semnificative din punct de vedere evolutiv (ESU). Fluxul de gene inter-descendență este indicat pentru speciile 1 și 3. În acest caz, independența evolutivă poate fi tranzitorie, deoarece două linii aparținând aceleiași specii nu sunt izolate reproductiv. În timp ce rămân ca linii distincte (specii), hibridizarea interspecifică poate apărea cu schimb de gene limitat (schimbare de culoare) între specii.
implementarea unor strategii eficiente de conservare care vizează protejarea atât a speciilor, cât și a diversității genetice ar beneficia atât de o viziune mai consistentă a modului în care delimităm speciile și alte unități în continuumul populație-specie, cât și de modul în care aceste unități sunt interpretate în diferite legislații la nivel global. Noile capabilități de la genomics oferă atât oportunități, cât și provocări pentru atingerea acestor obiective. În cele ce urmează considerăm: (i) modul în care analizele la scară genomică pot fi reconciliate cu o abordare conservatoare a delimitării și denumirii speciilor, identificând în același timp componentele majore ale diversității genetice în cadrul speciilor, (ii) modul în care legislația de conservare existentă pe mai multe continente, care definește „speciile” pentru a include o gamă largă de categorii infra-specifice, poate fi utilizată având în vedere rezoluția înaltă oferită de ecranele genomului și (iii) modul în care perspectivele asupra proceselor de speciație și structurarea variației de la genomică se aliniază legislației și apoi în management. Apoi, propunem câteva căi de urmat în alinierea unităților de conservare cu abordările legislative pentru a proteja întregul spectru al diversității de la populații prin specii.
genomica, speciația și taxonomia
ca multe domenii ale științei biologice, sistematica, practica numirii taxonilor și înțelegerea relațiilor lor și a modului în care se formează, este semnificativ avansată de Genomică și modele statistice. Fie prin secvențierea întregului genom (de exemplu, Nater și colab., 2017) sau eșantionarea genomului (SNPs, captură țintă-secvențiere; Jones și Good, 2016; Leache și Oaks, 2017), Avem acum o rezoluție fără precedent a modelelor de diversitate genetică (Funk și colab., 2012). Cuplate cu modele statistice din ce în ce mai puternice pentru deducerea Istoriilor divergenței genomice și a speciilor, aceste date oferă noi perspective asupra proceselor evolutive care generează specii și variații genetice.aplicarea acestor noi instrumente generează două perspective opuse. Pe de o parte, multe studii dezvăluie populații genetice extrem de divergente în cadrul speciilor numite, așa-numitele specii criptice (Bickford și colab., 2007; Jorger și Schr, 2013; Struck și colab., 2017). Acest lucru nu este nimic nou, deoarece se bazează pe o lungă istorie a geneticii populației spațiale și a filogeografiei moleculare care a fost un efort productiv de trei decenii (Avise și colab., 1987). Ceea ce aduce genomica acestui efort este atât rezoluția crescută a structurii populației, cât și datele care sunt suficiente pentru a deduce statistic istoriile populației și pentru a testa modele alternative de divergență (Degnan și Rosenberg, 2009; Sukumaran și Knowles, 2017). Pe de altă parte, analizele la scară genomică relevă, de asemenea, că (așa cum a fost recunoscut de mult timp pentru plante), schimbul genetic (introgresia) între speciile de animale este mai frecvent decât se credea anterior, atât în timpul speciației, cât și după aceasta, și poate conduce chiar noi radiații adaptive (Mallet, 2007; Rieseberg și Willis, 2007; Arnold și Kunte, 2017) Figura 1. Împreună, aceste perspective subliniază în continuare că speciația este prelungită (Roux și colab., 2016) și dezvăluie forțele opuse care operează prin continuumul filogeografie-filogenetică (Edwards și colab., 2016).
cum se leagă toate acestea de obiectivele de taxonomie și conservare? Există o îngrijorare puternică și justificată că rezoluția crescută oferită de ecranele genomului ar putea duce la o supra-divizare a speciilor (Isaac și colab., 2004), potențial restricționând flexibilitatea managementului și expediind populații mici divergente genetic la consangvinizare și eventuală Dispariție (Franham și colab., 2012). În schimb, Gippoliti și colab. (2018) susțin că inerția taxonomică, care a dus la eșecul anterior de a recunoaște multe specii de ungulate africane, a dus la o depresie de creștere în urma amestecării și la eșecul de a proteja taxonii restricționați geografic. Punctul lor de vedere (și a se vedea, de asemenea, Morrison și colab., 2009) este că nu există dovezi că inflația taxonomică are efecte negative asupra conservării ungulatelor (Gippoliti și colab., 2018). O preocupare conexă este că taxonomia instabilă întârzie atât practica conservării, cât și percepțiile publice (Garnett și Christidis, 2017). Aceste preocupări sunt în parte determinate de dezbaterea de lungă durată în jurul conceptelor de specii și a metodelor de diagnostic în biologia sistematică, în special aplicarea conceptului de specii filogenetice care susține că speciile sunt unități minim diagnosticabile (Cracraft, 1983). Cu toate acestea, majoritatea taxonomilor sunt în mod inerent conservatori, numind specii doar atunci când există concordanță pe mai multe linii de dovezi (Dayrat, 2005; Padial și colab., 2010; Schlick-Steiner și colab., 2010: Yeates și colab., 2011). Opinia noastră este că speciile ar trebui să reprezinte descendențe robuste în evoluție independentă, care rămân în mare parte intacte atunci când sunt în contact cu rudele apropiate—în concordanță cu intenția conceptului de specie evolutivă (Wiley, 1978) și a conceptului de specie generalizată (de Queiroz, 1998, 2007). În practică, acest lucru necesită dovezi directe ale izolării reproductive sau surogate rezonabile ale acestora, permițând în același timp un schimb genetic limitat (Singhal și colab., 2018).
datele genomice oferă o putere mai mare decât înainte pentru a identifica diferențierea și divergența în interiorul și între specii, necesitând o analiză mai mare a modului în care aceasta afectează delimitarea unităților taxonomice. În cazul în care populațiile sunt atât puternic divergente genetic, cât și fenotipic în raport cu speciile deja numite, există un caz puternic pentru denumirea acestora ca specii separate. Dacă populațiile sunt fenotipice puternic divergente, cu o separare genomică redusă, atunci există un apel de judecată dacă aceste diferențe fenotipice sunt (i) ereditare și (ii) susceptibile de a provoca izolare reproductivă substanțială. Scanările genomului pot rezolva astfel de situații prin testarea fluxului genetic restricționat. Problema mai dificilă este în cazul în care sunt detectate populații similare morfologic, dar divergente genetic. Astfel de populații pot fi ușor diagnosticate ca linii evolutive separate prin aplicarea metodelor de delimitare Statistică la datele multilocus (Carstens și colab., 2013; Rannala, 2015), dar sunt populații efemere sau specii durabile (Sukumaran și Knowles, 2017)? Cu siguranță este posibil, în cadrul regulilor nomenclaturii, să recunoaștem speciile numai din secvențele ADN, oferind diagnostice bazate pe secvențe (Tautz și colab., 2003; Cook și colab., 2010). De exemplu, Murphy și colab. (2015) delimitați și descrieți șase specii de amfipode de primăvară din deșertul Australian pe baza datelor secvenței ADN, în ciuda diferențelor morfologice clare. Cu toate acestea, acesta este un domeniu general de îmbunătățire în care sistematicii trebuie să aplice termenul „specie criptică” cu o mai mare consistență și rigoare (Struck și colab., 2017).
în situațiile în care datele secvenței ADN oferă o caracteristică cheie potențială definitorie pentru recunoașterea unei specii, sugerăm că este deosebit de important să se aplice alte linii de dovezi pentru a evita suprapunerea (Oliver și colab., 2015). O abordare este de a testa izolarea reproductivă substanțială în cazul în care astfel de populații formează zone naturale de contact. De exemplu, într-o analiză genomică comparativă a zonelor de contact dintre perechile de descendență ale șopârlelor Carlia care nu se disting fenotipic, indicii de izolare genetică au fost puternic corelați cu timpul de divergență (Singhal și Moritz, 2013; Singhal și Bi, 2017) (Figura 2). Cele mai divergente dintre aceste perechi de descendență au fost acum recunoscute formal ca specii separate, în timp ce descendențele mai strâns legate (dar diagnosticabile statistic) cu dovezi ale schimbului genetic extins genomic între zonele de contact nu au fost (Singhal și colab., 2018). Având în vedere relația empirică puternică dintre divergență și izolarea reproductivă la acești taxoni, acești autori au separat în continuare două izolate alopatrice și criptice (ale unei specii strâns legate) care aveau o divergență genomică chiar mai mare decât cele delimitate mai sus. Extinderea acestei izolații reproductive metrice informate la alte specii congenerice cu descendențe filogeografice profund divergente, dar similare morfologic (Potter și colab., 2016, 2018; Afonso Silva și colab., 2017a) sugerează că mai multe specii rămân de descris, în special pe insule, (Figura 2, Carlai Amax și C. rufilatus) și, într-adevăr, unele astfel de specii au fost descrise în urma analizei fenotipice complementare (Afonso Silva și colab., 2017b). Alte linii cu divergențe mai recente, dar încă delimitate statistic ca linii separate folosind un set mare de gene nucleare, pot fi reprezentate ca unități semnificative evolutiv (ESU) în cadrul speciilor.
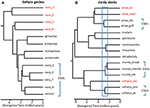
Figura 2. Rezoluția filogenomică a limitelor speciilor la șopârlele din tropicele musonice australiene (Amt). În fiecare caz, relațiile și timpii de divergență relativă sunt estimați de la 100 de exoni folosind coalescența multispecifică implementată în StarBEAST2, dar scalele de timp dintre cele două exemple nu sunt neapărat aceleași. (A) Rezultate pentru grupul nana de gehyra geckos (Moritz și colab., 2018). Diversitatea filogenetică a descendențelor din cadrul G. nana, așa cum a fost recunoscută anterior, a inclus alte patru specii deja descrise și distincte morfologic. Analizele ulterioare ale suprapunerilor și morfologiei intervalului au condus la recunoașterea fiecăreia dintre cele mai divergente linii filogenetice (etichete cu taxon roșu) ca specii separate (Doughty și colab., 2018). Liniile (inseparabile morfologic) rămase în G. nana așa cum sunt recunoscute acum pot fi considerate ESU. (B) rezolvarea continuă a limitelor speciilor din carlia skinks din AMT. Unele linii au fost deja recunoscute ca specii pe baza unei combinații de delimitare genetică a speciilor și identificarea post-hoc a diferențelor morfologice diagnostice (C. insularis și C. isostriacantha; Afonso Silva și colab., 2017b). Pentru alte complexe (Potter și colab., 2016, 2018), revizuirile taxonomice sunt în curs. Dreptunghiul albastru indică intervalul timpilor de divergență în care s-a observat o izolare reproductivă substanțială, corelată cu divergența, în zonele de contact dintre descendențele criptice morfologic ale speciilor înrudite (Singhal și Bi, 2017). Conservator și următor (Singhal și colab., 2018), recunoaștem ca specie candidată trei linii genetice cu divergențe mai mari decât acest prag empiric. Acest lucru ar lăsa ESU-uri distincte genetic în fiecare dintre Carlia Amax, Carlia munda și Carlia rufilatus (paranteze albastre). Barele de scară reprezintă estimări ale timpului de divergență în milioane de ani.
în cazuri extreme, metodele genomice dezvăluie că speciile taxonomice unice nu sunt doar heterogene genetic, ci, într-un sens filogenetic, conțin și alte specii numite. Un astfel de” parafil ” poate apărea prin speciația recentă dintr-o singură specie subdivizată genetic (Patton și Smith, 1994), dar generează și ipoteza că taxonul eterogen genetic poate reprezenta mai multe specii. Din nou, sunt necesare mai multe dovezi. De exemplu, analiza filogenomică a unui complex de șopârle Gehyra gecko din nordul Australiei a delimitat nouă linii evolutive, cu patru specii deja numite filogenetic încorporate printre ele (Moritz și colab., 2018; Figura 2). Dovezile ulterioare ale suprapunerii geografice (păstrând în același timp separarea filogenetică) și diferențele morfologice subtile au susținut denumirea celor patru linii cele mai divergente filogenetic (Doughty și colab., 2018). În schimb, liniile mai strâns legate (dar separabile statistic) au avut distribuții adiacente mai degrabă decât suprapuse, dar nu au fost diagnosticabile morfologic și, în consecință, au fost păstrate ca o singură specie (G. nana sensu stricto), deși cu ESU multiple, în așteptarea unor investigații suplimentare (Figura 2). Scanările ulterioare ale genomului probelor peste granițele acestor ESU au arătat că cel puțin unele au un schimb genetic substanțial, validând abordarea originală și conservatoare adoptată pentru recunoașterea speciilor.
în alte cazuri, studiile genomice ale plantelor, chiar și prin simpla ordonare a SNP-urilor, pot dezvălui divergențe puternice și neprevăzute între eșantioanele populației, sugerând prezența taxonilor criptici care pot fi apoi analizați pentru caracterele fenotipice diagnostice. De exemplu, dovezile bazate pe SNP pentru divergența genomică între populații și diferențele ecotipice în Eucalyptus salubris au evidențiat două linii moleculare distincte care și-au menținut caracterul distinctiv într-o zonă de suprapunere geografică, adaptarea ecotipică fiind considerată a fi un factor important în minimizarea fluxului de gene între linii. (Steane și colab., 2015; Figura 3). Într-un alt caz, în pelargoniul Australian, analiza genomică, combinată cu studii morfologice și reproductive, a dus la schimbări semnificative în taxonomie, atât divizarea, cât și aglomerarea taxonilor, în special în cazul în care trăsăturile morfologice utilizate anterior se dovedesc a fi labile și discordante cu filogenia (Nicotra și colab., 2016).
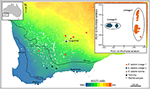
Figura 3. Analiza genomică utilizând datele SNP a identificat linii criptice în Eucaliptul larg răspândit Salubris. Localizarea populațiilor de eșantioane din fiecare descendență este prezentată într-un gradient al indicelui de ariditate în sud-vestul Australiei de vest. PCoA bazat pe distanțe genetice binare derivate din 16.122 markeri DArTseq, prezintă două linii distincte. Adaptat de la Steane și colab. (2015).
cazurile de mai sus ilustrează abordarea progresivă de testare a ipotezelor care a fost utilizată în taxonomia Integrativă (Padial și colab., 2010; Yeates și colab., 2011). Mai mult, aceste cazuri folosesc o formă de secvențiere a reducerii genomului pentru a obține date de înaltă calitate din 1000 de gene independente, oferind o încredere mult mai mare în rezultate decât este cazul folosind doar plastide loci sau câteva gene nucleare. Un exemplu al modului în care aplicarea taxonomiei integrative poate modifica dramatic percepțiile asupra diversității provine dintr-un studiu combinat genetic, acustic și morfologic al broaștelor din Madagascar, care aproape a dublat diversitatea speciilor cunoscute (Vieites și colab., 2009). Acești autori au propus o serie de termeni care reflectă starea rezoluției și congruenței taxonomice: Specii Candidate confirmate, pentru taxoni Divergenți genetic și fenotipic, dar care nu au fost încă numiți; specii Candidate neconfirmate pentru taxoni Divergenți genetic în care morfologia și acustica nu fuseseră încă examinate; și linii profund Conspecifice pentru entități cunoscute a fi divergente genetic, dar criptice fenotipic. Analizele genomice nu sunt încă aplicate acestor taxoni, dar cu siguranță ar informa deciziile cu privire la descendențele conspecifice față de speciile candidate.
abordările genomice, chiar și atunci când sunt aplicate conservator, vor duce fără îndoială la divizarea în continuare a taxonilor conservatori morfologic. Dar, dimpotrivă, există exemple clare în care aceleași abordări dezvăluie specii supra-divizate sau diagnosticate incorect (Nicotra și colab., 2016). Desigur, există deja multe studii moleculare asupra complexelor de specii de plante și animale bazate pe seturi de date la scară mai mică (de ex., microsatelit, ADNmt și ADNmt) care au identificat unde speciile au fost diagnosticate greșit sau unde mai multe specii au fost circumscrise taxonomic, dar sunt acum considerate a fi o singură specie. De exemplu, Rabosky și colab. (2014) au folosit filogeniile genei mitocondriale și nucleare pentru a revizui complexul scincid Ctenotus inornatus, rezultând o reducere netă a speciilor și realinierea limitelor speciilor; trăsăturile morfologice aplicate anterior, deși convenabile pentru biologii de teren, au fost foarte labile și în mare parte nu au reușit să se potrivească cu limitele speciilor diagnosticate genetic. La plante, unele dintre cele mai cunoscute exemple se găsesc în orhidee, cum ar fi genul orhideelor Europene Ophrys și genul orhideelor australiene Caladenia, unde studiile moleculare indică semnificativ mai puține entități care ar fi recunoscute ca specii, decât cele bazate pe trăsături morfologice și sindroame de polenizare pseudocopulatorie (vezi Devey și colab., 2008; Swarts și colab., 2014).
datele la scară genomică, atunci când sunt interpretate în mod conservator și ideal cu analize paralele ale fenotipurilor, vor continua să conducă la modificări taxonomice, în special la taxonii conservatori morfologic (fi Inquier et al., 2018). Dar, spre deosebire de preocupările exprimate de unii (Garnett și Christidis, 2017), considerăm că aceasta este urmărirea corectă a sistematicii evolutive, chiar dacă rezultatele sunt temporar incomode.
genomica și unitățile de conservare intraspecifice
deși mai puțin controversate decât delimitarea taxonomică și dezbaterea la nivel de specie, dar în multe privințe la fel de complexe, a fost discuția semnificativă asupra definirii și delimitării unităților de conservare ca elemente importante ale diversității intraspecifice care trebuie luate în considerare în acțiunile de conservare (Ryder, 1986; Waples, 1991; Dizon și colab., 1992; Moritz, 1994; Vogler și de Salle, 1994; Crandall și colab., 2000; Fraser și Bernatchez, 2001; Avise, 2004; Funk și colab., 2012) (Figura 4). Având în vedere abordarea prudentă a recunoașterii speciilor din dovezile genomice pe care le susținem mai sus (și vezi și Oliver și colab., 2015; Singhal și colab., 2018), este inevitabil ca unele specii taxonomice să conțină în continuare populații izolate de mult timp în interiorul lor. Concepte precum unități semnificative din punct de vedere evolutiv (ESU) sau Unități desemnabile (DUs) au fost propuse pentru a completa taxonomia existentă (Ryder, 1986; Waples, 1991; Moritz, 1994; Mee și colab., 2015). Ca și în cazul speciilor, interpretările și criteriile variază, dar intenția comună este de a recunoaște elementele majore ale diversității intraspecifice pentru acțiunile de conservare (Funk și colab., 2012). Moritz (1994, 2002) a susținut o abordare moleculară simplistă, dar practică, pentru identificarea segmentelor care evoluează independent în cadrul speciilor, axa „istorică” a diversității; în timp ce alții au pus mai mult accent pe axa „adaptivă” pentru a include diversitatea funcțională (Ryder, 1986; Crandall și colab., 2000; Fraser și Bernatchez, 2001; Waples și colab., 2001; Mee și colab., 2015) (Figura 4). Oricare ar fi meritele relative ale acestor alternative, este clar că instrumentele tehnice și analitice ale genomicii populației pot identifica acum solid ambele componente ale diversității intraspecifice (Funk și colab., 2012; Hoban și colab., 2013; Whitlock, 2014; Yang și Rannala, 2014).
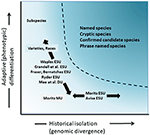
Figura 4. Separarea diversității genetice în două componente: variație adaptivă care apare prin selecție naturală și testată prin analiza fenotipurilor și izolarea istorică sau divergența neutră evaluată prin divergența genomică. Zona umbrită mai întunecată de deasupra curbei indică condițiile în care populațiile sunt susceptibile de a fi considerate specii separate în majoritatea conceptelor. Sunt prezentate diferite concepte de unități de conservare și categorii taxonomice infra-specifice în raport cu cele două axe ale diversității genetice (după Moritz, 2002).
aceasta duce la întrebarea: de ce să nu numim ESUs doar subspecii? Subspeciile au fost în mod tradițional definite și privite atât de botaniști, cât și de zoologi ca seturi de populații distincte fenotipic, alopatrice, care se pot intergrada între ele la granițele geografice (Mayr, 1942; Grant, 1981). Ca atare, ele reflectă potențial divergența adaptivă fără a fi neapărat izolate istoric. Denumirile subspeciilor sunt utilizate pe scară largă în plante, cu diferențe geografice și ecologice principalele mijloace prin care se disting (Hamilton și Reichard, 1992). În măsura în care subspeciile manifestă adaptare locală, cu sau fără flux genetic continuu, sugerăm că subspeciile definite fenotipic ar trebui să continue să fie recunoscute în conservare și să nu fie combinate cu ESU, cel puțin nu ESU așa cum sunt definite pentru a reprezenta populații izolate istoric.în același mod în care genomica populației oferă o precizie mult mai mare în identificarea ESU-urilor prin utilizarea atât a locilor neutri, cât și a celor adaptivi, oferă, de asemenea, un mijloc de utilizare a unui număr mare de loci neutri pentru a delimita mai ușor unitățile de Management (Mu) ca unități demografic independente caracterizate de fluxul genetic restricționat (Funk și colab., 2012). În timp ce cea mai mare parte a concentrării noastre aici este pe unitățile ESU și analoge, cum ar fi DUs, care pot fi recunoscute ca fiind principalele unități intra-specifice pentru conservare și au fost acceptate pentru protecție în unele jurisdicții, Mu au fost, de asemenea, identificate reprezentând seturi de populații independente demografic (vezi Palsb Okticll și colab., 2007) care poate fi reușit să păstreze ESU mai mare (Moritz, 1994; Funk și colab., 2012). Unitățile de gestionare pot fi importante în monitorizarea și gestionarea populațiilor de specii și au fost denumite și „stocuri”, deosebit de relevante pentru gestionarea pescuitului (Avise, 2004; Laikre și colab., 2005). Definiția MUs de Moritz (1994) ca „populații cu divergență semnificativă a frecvențelor alelelor la loci nucleari sau mitocondriali, indiferent de caracterul distinctiv filogenetic al alelelor” este în general acceptată pe scară largă, deși, așa cum a subliniat Palsb Okticll și colab. (2007), conectivitatea demografică este factorul cheie nu nivelul fluxului genetic istoric.
delimitarea unităților de conservare ca elemente importante ale diversității intraspecifice are, de asemenea, implicații semnificative pentru salvarea genetică a populațiilor consangvinizate mici prin restabilirea fluxului genetic sau prin augmentare și devine din ce în ce mai clar că salvarea genetică trebuie luată în considerare mai larg dacă se va evita dispariția crescută a populației (Love Stowell și colab., 2017; Ralls și colab., 2018). A fost propus de Moritz (1999) că amestecarea Mu-urilor, dar nu a ESU-urilor, ar putea fi considerată o strategie adecvată pentru salvarea genetică. După cum a subliniat Frankham și colab. (2012) amestecarea speciilor recunoscute taxonomic nu ar fi, în general, considerată acceptabilă de către manageri, caz în care supraexplitarea taxonomică ar putea restricționa oportunitățile de salvare a populațiilor consangvinizate mici. Sugerăm că, deși o abordare mai riguroasă a delimitării unităților intraspecifice, cum ar fi MUs, ar putea contribui la atenuarea acestei probleme, este, de asemenea, clar că estimarea genomică a ratelor de migrație și a divergenței adaptive are potențialul de a oferi îndrumări mult mai robuste nu numai în definirea limitelor speciilor și a variației intraspecifice, ci și în proiectarea strategiilor de salvare genetică.
Filogenomica și conservarea
genomica combinată cu metodele filogenetice poate rezolva cu ușurință relațiile și distanța evolutivă între taxoni, precum și delimitarea speciilor. Acest lucru se realizează cel mai bine folosind metode de arbori de specii, mai degrabă decât concatenare, deoarece prima explică eterogenitatea arborelui genic și evită supra-inflația lungimilor vârfurilor datorită polimorfismului ancestral (Edwards și colab., 2016; Ogilvie și colab., 2016). În timp ce numărul masiv de loci nu este adesea esențial, includerea mai multor loci îmbunătățește adesea rezoluția atât a relațiilor, cât și a lungimilor ramurilor (de exemplu, Blom și colab., 2016). Importanța conservării este dublă. În primul rând, evaluările filogenetice permit luarea în considerare a caracterului distinctiv al speciilor sau ESUs, nu doar dacă sunt descrise. Astfel, programul EDGE (Isaac și colab., 2007) pledează pentru ponderarea speciilor pentru conservare pe baza cantității de istorie evolutivă unică pe care o reprezintă, pe lângă criteriile obișnuite ale IUCN. În al doilea rând, lungimile ramurilor în sine pot reprezenta valori ale biodiversității, independente de taxonomie (Faith, 1992), care și-a demonstrat valoarea în prioritizarea zonelor pentru conservare (laici și colab., 2015; Rosauer și colab., 2016, 2018). În acest sens, adoptarea metricilor filogenetice folosind măsuri precum diversitatea filogenetică și endemismul filogenetic face posibilă evidențierea zonelor cu o diversitate evolutivă ridicată care pot fi vizate pentru protecție și li se acordă o prioritate ridicată în orice proces de planificare pentru îmbunătățirea conservării biodiversității. Această abordare poate spori protecția diversității genetice și a proceselor evolutive pentru diverse grupuri de organisme într-o serie de scale geografice (laici și colab., 2015) luarea deciziilor cu privire la zone pentru a acorda prioritate mai robustă capriciilor modului în care dovezile genomice sunt traduse în recunoașterea speciilor sau a unităților intra specifice.
lumea reală—definiții Legislative
Politica și cadrele legislative pentru conservarea biodiversității variază de la convențiile și strategiile internaționale, până la strategiile și legislația națională și legislația de stat sau regională și strategiile din cadrul țărilor. Principalele instrumente internaționale privind conservarea biologică includ Convenția privind diversitatea biologică implementată prin Convenția părților (COP), Convenția Patrimoniului Mondial, Strategia Globală pentru biodiversitate, procesul de la Montreal pentru conservarea pădurilor, Convenția Ramsar privind Zonele umede, Convenția privind speciile migratoare, Convenția privind comerțul internațional cu specii pe cale de dispariție (CITES), Lista Roșie a Uniunii Internaționale pentru Conservarea Naturii (IUCN) privind speciile amenințate și Convenția-cadru a Organizației Națiunilor Unite privind schimbările climatice. Toate aceste charte întruchipează principiul conservării speciilor și a diversității genetice ca obiectiv de bază al conservării biodiversității, speciile fiind unitatea definită în general pentru protecție. Cu toate acestea, definițiile speciilor în conformitate cu legislația și directivele internaționale sunt extrem de variabile. Acestea încorporează recunoașterea taxonomică a speciilor și a taxonilor intraspecifici, dar pot încorpora și concepte genetice/evolutive care sunt adesea acoperite de definiția legală a unei „specii” (Tabelul 1).

Tabelul 1. Legislația și directivele mondiale care definesc „speciile” pentru listare și protecție.având în vedere gama largă de niveluri infra-specifice care pot fi protejate (Tabelul 1), legislația și procesele de listare asociate ale țărilor ar trebui să faciliteze conservarea diversității genetice. De exemplu, definiția unei „specii”, atunci când se ia în considerare legislația din întreaga lume (Tabelul 1), poate include: specii, subspecii, soiuri și populații geografice și/sau genetice distincte, în timp ce în conformitate cu legislația sud-africană poate include și soiuri, rase geografice, tulpini și hibrizi. Astfel, în majoritatea legislației la nivel global, o „specie” include entități care reflectă diferite niveluri infra-specifice care fac parte dintr-o distribuție mai largă a speciilor și reflectă în grade diferite continuumul populație-specie. Aceasta este paralelă cu dificultatea inevitabilă cu care se confruntă taxonomiștii, biologii evolutivi și geneticienii de conservare în găsirea unui mod mai consistent și mai uniform de a specifica unitățile din acest continuum.
unii susțin că preocupările legate de implicațiile politice ale speciilor și incertitudinea infra-specifică sunt deplasate (Hey și colab., 2003), dar suntem încă de părere că, deși legislația poate oferi un mecanism de protejare a diversității genetice la o serie de niveluri, sarcina biologilor este de a oferi mai multă claritate și consecvență cu privire la modul în care aceste niveluri ar trebui delimitate de la specii până la populații individuale. În prezent, managerii și legislatorii rămân cu o gamă uluitoare de concepte de specii și unități infra-specifice furnizate de oamenii de știință atunci când delimitează unități în continuumul populație-specie. De exemplu, toate jurisdicțiile consideră speciile recunoscute de taxonomiști, ca unități cheie pentru conservare (Tabelul 1), totuși există încă o dezbatere largă între taxonomiști în jurul conceptelor speciilor și modul în care speciile sunt diagnosticate în practică variază în interiorul și între grupuri. Acest lucru a dus la un apel recent pentru o abordare mai reglementată a taxonomiei (Garnett și Christidis, 2017), la care alții au contracarat că reglementarea taxonomiei nu este răspunsul (Thomson și colab., 2018). Deși suntem de acord cu unele dintre punctele făcute de Thomson și colab. (2018) sugerăm că este destul de naiv să propunem că legislația în sine trebuie să se schimbe pentru a face referire în mod explicit la conceptul specific de taxon implicat de un nume. Acestea și dezbaterile anterioare evidențiază necesitatea ca biologii și taxonomii evoluționiști să preia conducerea în asigurarea clarității (transparenței) în modul în care traduc dovezile genomice și de altă natură la delimitarea speciilor sau a unităților intraspecifice și modul în care acești taxoni se raportează la legislația relevantă pentru conservare și protecție.
traducere către Management
după cum sa discutat mai sus, toate legislațiile și directivele recunosc nivelul speciilor ca unitate fundamentală de conservare, dar prevăd, de asemenea, recunoașterea diferitelor unități infra-specifice. Mulți au, de asemenea, o cerință implicită pentru denumirea formală și descrierea taxonomică a speciilor atunci când sunt luate în considerare pentru prioritizarea conservării și protecția formală. De exemplu, în conformitate cu liniile directoare privind listarea Roșie a IUCN, listarea speciilor fără nume este descurajată și există cerința ca noua descriere a speciilor să fie publicată în termen de patru ani de la includerea speciilor pe Lista Roșie a IUCN sau să fie eliminată. Acest lucru evidențiază importanța denumirii și descrierii taxonomice a entităților, atât la nivel de specii, cât și la nivel infra-specific, informațiile taxonomice limitate pentru multe grupuri de plante și animale (Mace, 2004; Oliver și Lee, 2010; Wilson, 2017) și necesitatea de a accelera efortul taxonomic, în special în zonele bogate în biodiversitate (Riedel și colab., 2013; Wege și colab., 2015; Wilson, 2017).
speciile taxonomice și categoriile taxonomice intraspecifice, cum ar fi subspeciile și soiurile, oferă în general unitățile de bază pentru listarea formală, prioritizarea și protecția conservării, totuși, în unele jurisdicții, alte categorii de unități de conservare intraspecifice sunt recunoscute formal. Acestea sunt în general identificate folosind date genomice și fenotipice combinate. De exemplu, Legea privind speciile pe cale de dispariție din SUA recunoaște segmente distincte de populație (DPSs) care se bazează în general pe ESU (Waples, 1991), în timp ce legea canadiană privind speciile în pericol recunoaște DUs. O interpretare operațională a” populațiilor distincte ” în conformitate cu legea canadiană privind speciile la risc, DUs își propune să identifice seturi discrete de populații care reprezintă componente importante ale moștenirii evolutive a speciilor și care este puțin probabil să fie înlocuite prin dispersie naturală (Mee și colab., 2015).
atât DUs, cât și DPSs se suprapun conceptual cu ESU (Waples și colab., 2013) și au fost aplicate cel mai mult peștilor din America de Nord, inclusiv salmonidelor din SUA de Nord-Vest (Waples, 1995) și Coregonidelor canadiene (Mee și colab., 2015) interesant DPSs în SUA se aplică numai vertebratelor, în timp ce în Canada DUs poate fi aplicat oricărui organism care poate fi listat în conformitate cu Legea sa (vezi Waples și colab., 2013), cu DUs listate în prezent pentru artropode, moluște și plante. În Australia, cum ar fi Canada, populații distincte pot fi listate pentru orice organism recunoscut în temeiul Legea privind protecția mediului și conservarea biodiversității din 1999 (EPBC Act), deși nu sunt o categorie denumită în mod specific, cum ar fi DUSs și DPSs. Cu toate acestea, până în prezent au fost recunoscute doar pentru vertebrate. În SUA există 127 DPSS vertebrate enumerate pentru conservare (https://ecos.fws.gov/ecp/); în Canada au fost enumerate 70 DUs care acoperă toate organismele (http://www.registrelep-sararegistry.gc.ca/species/schedules_e.cfm?id=1); și în Australia populațiile de 7 specii de vertebrate care acoperă mamifere, pești și păsări au fost recunoscute ca având o stare de conservare specifică în conformitate cu legea EPBC (http://www.environment.gov.au/cgi-bin/sprat/public/publicthreatenedlist.pl). În timp ce evaluările acestor populații din SUA și Canada utilizează în general informații genetice (Waples și colab., 2013), în Australia se bazează de obicei pe identificarea geografică și jurisdicțională, deși în unele dintre aceste cazuri au fost susținute de analiza diferențierii genetice pentru mamifere (Firestone și colab., 1999; Armstrong, 2006) și fish (Colgan și Paxton, 1997; Ahonen și colab., 2009).lista unităților de conservare infra-specifice pentru plante și nevertebrate din Canada (DUs), dar nu în SUA sau Australia, evidențiază o altă problemă care nu este legată în mod specific de delimitarea unităților de conservare, ci mai larg de inconsecvențele semnificative în prioritizarea conservării biodiversității; vertebratele iconice, cum ar fi păsările, mamiferele și peștii, sunt un accent în multe jurisdicții, în timp ce alte organisme nu sunt. Această listă preferențială a populațiilor anumitor organisme reflectă o serie de probleme care au mai puțin de-a face cu știința delimitării unităților de conservare și mai mult de-a face cu listarea populațiilor bazate pe conștientizarea publicului, oportunitatea politică și natura carismatică a organismului care urmează să fie listat (vezi Waples și colab., 2013).în timp ce în Australia unele populații de mamifere, păsări și pești bine cunoscuți sunt protejate în temeiul Legii EPBC, există acum un corp semnificativ de informații bazate pe studii moleculare în reptile, nevertebrate și plante în care populațiile pot fi recunoscute ca ESUs sau MUs, iar multe dintre acestea au o semnificație de conservare, având în vedere gama lor geografică îngustă, dimensiunea redusă a populației, rata de declin și susceptibilitatea la procese amenințătoare. De exemplu, Exemple izbitoare de populații foarte structurate și linii profund divergente în cadrul speciilor care s-ar potrivi cu ușurință cerinței unui ESU pot fi găsite într-o serie de vertebrate cu dispersie redusă de la tropice (Figura 2) și în altă parte. În mod similar, în nevertebratele australiene, cum ar fi amfipodele și milipedele, structura genetică semnificativă și modelele filogeografice susțin delimitarea grupurilor de populație discrete ca ESU care sunt disparate geografic, cu intervale geografice înguste, limitate la habitate specifice și reprezentate de doar câteva populații (Finston și colab., 2007; Murphy și colab., 2013; Nistelberger și colab., 2014a). Adesea, aceste endemice cu rază scurtă de acțiune sunt taxoni de preocupare pentru conservare (Harvey și colab., 2011) și sunt co-localizate în hotspoturile endemismului (de exemplu, Oliver și colab., 2017), care sunt ele însele priorități înalte pentru atenția conservării multi-taxonilor (Rosauer și colab., 2018), cum ar fi regiunea Kimberley și gama Stirling din sud-vestul Australiei de Vest.
în plantele australiene există, de asemenea, un număr semnificativ de studii genetice și filogeografice ale populației care identifică grupuri distincte de populație dintr-o serie de specii din diferite genuri care reflectă sisteme de populație disjuncte și izolate istoric, complexități geologice și edafice și care ocupă habitate contrastante în ceea ce privește vegetația și clima (vezi Broadhurst și colab., 2017). Populațiile distincte genetic sunt deosebit de evidente la speciile din sud-vestul Australiei de Vest (Coates, 2000; Byrne și colab., 2014) cu o serie de studii care evidențiază o structură genetică semnificativă și linii extrem de divergente într-o serie de genuri și familii de plante, inclusiv: salcâm (Mimosaceae; Coates, 1988), Atriplex (Amaranthaceae; Shepherd și colab., 2015), Banksia (Proteaceae; Coates și colab., 2015), Lambertia (Proteaceae; Byrne și colab., 2002), Hakea (Proteaceae; Sampson și colab., 2014), Eremophila (Scrophulariaceae; Llorens și colab., 2015), Pultanaea (Fabaceae; Millar și Byrne, 2013) și Calothamnus (Myrtaceae; Nistelberger și colab., 2014b). În multe cazuri, aceste populații sunt recunoscute în planificarea conservării de către agențiile guvernamentale de stat și luate în considerare în evaluările impactului asupra mediului (vezi Shepherd și colab., 2015), chiar dacă nu sunt enumerate pentru protecție în temeiul legislației. Cu toate acestea, recunoaștem că recunoașterea formală a acestor unități ar îmbunătăți conservarea lor pe termen lung.
aceste exemple australiene evidențiază nevoia crescândă de a recunoaște acest strat de diversitate genetică sub nivelul speciilor. Provocarea este de a se asigura că populațiile foarte structurate și descendențele profund divergente care sunt de interes pentru conservare sunt protejate în fața cererilor concurente de atenție pentru conservare. Dincolo de speciile iconice, această provocare este evidentă și la nivel global, cu excepția Canadei. Eșecul de a proteja unitățile de conservare clar delimitate va crește probabil riscul de dispariție criptică și pierderea diversității genetice semnificative, care în multe cazuri poate implica descendențe și populații izolate istoric care au persistat în cadrul speciilor de milioane de ani (Moritz, 2002).
concluzii și recomandări
listarea speciilor și a diferitelor categorii infra-specifice în temeiul legislației privind speciile amenințate sau în cadrul organizațiilor internaționale, cum ar fi IUCN, indică un angajament de a proteja diversitatea genetică în cadrul continuumului populației speciilor. Cu toate acestea, abordările actuale ale conservării biodiversității se concentrează în mare măsură asupra sfârșitului speciilor acestui continuum, acordând mai puțină atenție diversității genetice și variației infra-specifice. Acest accent mai limitat pe variația genetică la nivel intra-specie sugerează că, în timp ce managerii de conservare recunosc din ce în ce mai mult valoarea luării în considerare în mod explicit a informațiilor genetice în acțiunile de management și prioritizare, adesea le lipsește informații acționabile de la oamenii de știință din domeniul biodiversității (Waples și colab., 2008; Laikre și colab., 2010; Cook și SGR, 2017; Taylor și colab., 2017). Acest decalaj între știință și practica conservării împiedică o abordare globală mai eficientă a conservării și gestionării diversității genetice.
sugerăm că lipsa de claritate din partea oamenilor de știință în domeniul biodiversității asupra unităților de conservare se bazează în mare parte pe inconsecvențe la două niveluri. Primul se referă la conceptele de specii multiple și la diferitele metode de diagnostic utilizate pentru delimitarea speciilor care creează inconsecvență între grupurile taxonomice. Al doilea se referă la o lipsă de coerență și acord de către taxonomi, biologi evoluționiști și biologi de conservare cu privire la modul de tratare a taxonilor infra-specifici și a altor entități, cum ar fi ESU, DUs și MUs.în timp ce considerăm că legislația este adesea deja suficient de flexibilă în ceea ce privește modul în care sunt definiți taxonii (sau „speciile”), susținem că este necesar să se abordeze sistematic continuumul speciilor populației și să se definească mai clar unitățile de conservare, cu o mai mare recunoaștere acordată conceptelor precum ESU și modul în care acestea se raportează la legislația relevantă. Subliniem că această problemă va deveni, fără îndoială, mai critică, având în vedere puterea genomicii de a detecta diferențierea la scări progresiv mai fine și utilizarea sa din ce în ce mai mare pentru a delimita speciile și entitățile infra-specifice.
Din analiza precedentă, oferim următoarele recomandări:
(i) datele genomice ar trebui interpretate în mod conservator la delimitarea speciilor, folosind o abordare taxonomică integrativă care include de obicei dovezi dincolo de datele genetice.
(ii) în cazul în care geneticienii identifică mai multe linii sau populații distincte în cadrul unei specii, iar acestea nu justifică ridicarea la nivelul speciilor și nu sunt recunoscute taxonomic, există un impuls mai puternic și o abordare mai uniformă pentru identificarea unităților intraspecifice clar justificate pentru evaluarea practică a stării de conservare sau a evaluărilor impactului și că acestea sunt recunoscute în mod explicit ca unități de conservare în contextul legislației relevante pentru jurisdicția respectivă.
(iii) IUCN, prin grupul său de specialiști în genetica conservării și subgrupurile sale regionale, ia în considerare elaborarea de orientări pentru recunoașterea și etichetarea unităților intraspecifice care se aliniază celor mai bune practici științifice.
(iv) viziunea mai nuanțată care apare din genomica evolutivă a speciilor și continuumul de la populații la specii ar trebui comunicată mai eficient de către oamenii de știință managerilor de conservare și factorilor de decizie politică.
în sens mai larg, facem ecou recomandărilor făcute recent într-un număr de recenzii (Frankham, 2010; Santamar OACA și m Oktoktndez, 2012; Hoban și colab., 2013; Moritz și Potter, 2013; laici și colab., 2015; Pierson și colab., 2016; Cook și SGR, 2017; Taylor și colab., 2017) care evidențiază nevoia oamenilor de știință de a îmbunătăți comunicarea și de a sensibiliza factorii de decizie politică, agențiile guvernamentale, organizațiile neguvernamentale și practicienii care rezultă din sistematica moleculară, taxonomia moleculară, filogeografia și cercetarea genetică a populației pot oferi informații critice despre conservarea diversității genetice și gestionarea proceselor evolutive.
contribuțiile autorului
DC, MB și CM au contribuit la pregătirea și revizuirea manuscrisului.
Declarație privind conflictul de interese
autorii declară că cercetarea a fost realizată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
mulțumiri
Acest proiect a fost susținut de Consiliul australian de cercetare (Grant arc Linkage nr. LP120200063), Universitatea Națională Australiană și Departamentul de biodiversitate, conservare și atracții din Australia de Vest.Afonso Silva, A. C., Bragg, J. G., Potter, S., Fernandes, C., Coelho, M. M. și Moritz, C. (2017a). Specialist Tropical versus generalist climatic: diversificarea și istoria demografică a speciilor surori de carlia skinks din nord-vestul Australiei. Mol. Ecol. 26:4045–4058. doi: 10.1111 / mec.14185
CrossRef Text Complet/Google Scholar
Afonso Silva, A. C., Santos, N., Ogilvie, H. A. și Moritz, C. (2017b). Validarea și descrierea a două noi skin-uri curcubeu din nord-vestul Australiei cu metode și morfologie coalescente multispecifice. PeerJ 5: e3724. doi: 10.7717 / peerj.3724
PubMed Abstract | CrossRef Full Text/Google Scholar
Ahonen, H., Harcourt, R. G. și Stow, A. J. (2009). ADN-ul Nuclear și mitocondrial dezvăluie izolarea populațiilor de rechini gri periclitați (Carcharias taurus). Mol. Ecol. 18, 4409–4421. doi: 10.1111 / j. 1365-294X.2009.04377.x
PubMed Abstract | CrossRef Full Text/Google Scholar
Armstrong, K. N. (2006). Structura filogeografică în Rhinonicteris aurantia (Chiroptera: Hipposideridae): implicații pentru conservare. Acta Chiropterologica 8, 63-81. doi: 10.3161/1733-5329(2006)82.0.CO
CrossRef Full Text | Google Scholar
Arnold, M. L., și Kunte, K. (2017). Schimb genetic adaptiv: o istorie încurcată a amestecului și a inovației evolutive. Tendințe Ecol. Evol. 32, 601–611. doi: 10.1016/j.copac.2017.05.007
PubMed Abstract | CrossRef Full Text/Google Scholar
Avise, J. C. (2004). Markeri moleculari, Istorie Naturală și Evoluție. A 2-a Edn. Sunderland, MA; Oxford: Oxford University Press.
Google Scholar
Avise, J. C., Arnold, J., Ball, R. M., Bermingham, E., miel, T., Neigel, J. E., și colab. (1987). Filogeografie intraspecifică: Podul ADN mitocondrial între genetica populației și sistematică. Ann. Rev. Ecol. Sys. 18, 489–522. doi: 10.1146 / annurev. es.18.110187.002421
CrossRef Full Text | Google Scholar
Bickford, D., Lohman, D. J., Sodhi, N. s, Ng, P. K., Meier, R., Winker, K., și colab. (2007). Specii criptice ca fereastră asupra diversității și conservării. Tendințe Ecol. Evol. 22, 148–155. doi: 10.1016/j.copac.2006.11.004
PubMed Abstract | CrossRef Full Text/Google Scholar
Blom, MPK, Bragg, JG, Potter, S. și Moritz, C. (2016). Contabilizarea incertitudinii în estimarea arborelui genic: rezumat-inferența arborelui speciilor coalescente într-o radiație provocatoare a șopârlelor australiene. Syst. Biol. 66, 352–366. doi: 10.1093/sysbio/syw089
PubMed Abstract | CrossRef Full Text/Google Scholar
Broadhurst, L., Breed, M., Lowe, A., Bragg, J., Catullo, R., Coates, D. J. și colab. (2017). Diversitatea genetică și structura florei australiene. Scafandri. Distrib. 23, 41–52. doi: 10.1111 / ddi.12505
CrossRef Full Text | Google Scholar
Byrne, M., Coates, D. J., Forest, F., Hopper, S. D., Krauss, S. L., Sniderman, J. M. K. și colab. (2014). „O floră diversă-specii și relații genetice”, în viața plantelor pe câmpiile de nisip din sud-vestul Australiei, un Hotspot global al biodiversității, editori H. Lambers (Crawley, WA: University of Western Australia Publishing), 81-99.
PubMed Abstract
Byrne, M., Macdonald, B. și Coates, D. J. (2002). Modele filogeografice și diversitate ridicată a adcpc în cadrul Acacia acuminata (Leguminosae:Mimosoideae). complex în Australia de Vest. J. Evol. Biol. 15, 576–587. doi: 10.1046 / j. 1420-9101.2002. 00429.x
CrossRef Full Text/Google Scholar
Carstens, B. C., Pelletier, T. A., Reid, N. M. și Satler, J. D. (2013). Cum să eșuezi la delimitarea speciilor. Mol. Ecol. 22, 4369–4383. doi: 10.1111 / mec.12413
PubMed Abstract | CrossRef Full Text/Google Scholar
Coates, D. J. (1988). Diversitatea genetică și structura genetică a populației în rarele zăbrele de iarbă Chittering, Acacia anomala Court. Aust. J. Bot. 36:273–286. doi: 10.1071 / BT9880273
CrossRef Text Complet/Google Scholar
Coates, DJ (2000). Definirea unităților de conservare într-o floră bogată și fragmentată: implicații pentru gestionarea resurselor genetice și a proceselor evolutive în plantele din Australia de sud-vest. Aust. J. Bot. 48, 329–339. doi: 10.1071/BT99018
CrossRef Full Text/Google Scholar
Coates, DJ, McArthur, SL, și Byrne, M. (2015). Pierderea semnificativă a diversității genetice în urma dispariției populației determinate de agenți patogeni în rara endemică Banksia brownii (Proteaceae). Biol. Conserv. 192, 353–360. doi: 10.1016 / j. biocon.2015.10.013
CrossRef Full Text/Google Scholar
Colgan, DJ, și Paxton, J. R. (1997). Genetica biochimică și recunoașterea unui stoc occidental de gemfish comun, Rexea solandri (Scombroidea: Gempylidae)., în Australia. Mar. Freshw. Res. 48, 103-118. doi: 10.1071 / MF96048
CrossRef Full Text/Google Scholar
Convenția privind diversitatea biologică (2007). Disponibil online la adresa:https://www.cbd.int/doc/meetings/cop-bureau/cop-bur-2007/cop-bur-2007-10-14-en.pdf
Cook, C. N., și SGR, C. M. (2017). Alinierea științei și a politicii pentru a realiza o conservare luminată evolutiv. Conserv. Biol. 31, 501–512. doi: 10.1111 / cobi.12863
PubMed Abstract | CrossRef Full Text/Google Scholar
Cook, L. G., Edwards, R. D., Crisp, MD și Hardy, N. B. (2010). Este necesar ca morfologia să fie întotdeauna necesară pentru descrierile speciilor noi? Inversează. Sys. 24, 322–332. doi: 10.1071 / IS10011
CrossRef Text Complet/Google Scholar
Cracraft, J. (1983). Concepte de specii și analiza speciației. Curr. Ornitol. 1, 159–187. doi: 10.1007/978-1-4615-6781-3_6
CrossRef Full Text | Google Scholar
Crandall, K. A., Bininda-Emonds, O. R., Mace, G. M. și Wayne, R. K. (2000). Luând în considerare procesele evolutive în biologia conservării. Tendințe Ecol. Evol. 15, 290–295. doi: 10.1016 / S0169-5347 (00)01876-0
PubMed Abstract | CrossRef Full Text | Google Scholar
Cutter, AD, și Payseur, Ba (2013). Semnături genomice de selecție la site-uri legate: unificarea disparității între specii. Nat. Părinte Genet. 14, 262–274. doi: 10.1038 / nrg3425
PubMed Abstract | CrossRef Full Text/Google Scholar
Dayrat, B. (2005). Către taxonomia Integrativă. Biol. J. Linn. Soc. 85, 407–415. doi: 10.1111 / j. 1095-8312.2005. 00503.x5
CrossRef Full Text/Google Scholar
de Queiroz, K. (1998). „Conceptul general de descendență a speciilor, criteriile speciilor și procesul de speciație”, în forme: specii și speciație, D. J. Howard,r S. H. Berlocher (New York, NY: Oxford University Press), 57-75.
Google Scholar
de Queiroz, K. (2007). Concepte de specii și delimitarea speciilor. Syst. Biol. 56, 879–886. doi: 10.1080 / 10635150701701083
PubMed Abstract | CrossRef Full Text/Google Scholar
Degnan, JH, și Rosenberg, N. A. (2009). Discordanța arborelui genic, inferența filogenetică și coalescența multispecifică. Tendințe Ecol. Evol. 24, 332–340. doi: 10.1016/j.copac.2009.01.009
PubMed Abstract | CrossRef Full Text/Google Scholar
Devey, D. S., Bateman, R. M., Fay, M. F. și Hawkins, J. A. (2008). Prieteni sau rude? Filogenetica și delimitarea speciilor în controversatul gen de orhidee European Ophrys. Ann. Bot. 101, 385–402. doi: 10.1093/aob/mcm299
PubMed Abstract | CrossRef Full Text/Google Scholar
Dizon, A. E., Lockyer, C, Perrin, W. F., Demaster, D. P. și Sisson, J. (1992). Regândirea conceptului de stoc – o abordare filogeografică. Conserv. Biol. 6, 24–36. doi: 10.1046 / j. 1523-1739.1992. 610024.x
CrossRef Full Text/Google Scholar
Doughty, P., Bourke, G., Tedeschi, L., Pratt, R., Oliver, P., Palmer, R. și colab. (2018). Delimitarea speciilor în Gehyra nana (Squamata: Gekkonidae). complex: evoluție morfologică criptică și divergentă în tropicele musonice australiene, cu descrierea a patru specii noi. Zootaxa, 4403: 201-244. doi: 10.11646/zootaxa.4403.2.1
PubMed Abstract | CrossRef Full Text/Google Scholar
Dynesius, M. și Jansson, R. (2014). Persistența liniilor din interiorul speciilor: un control neglijat al ratelor de speciație. Evoluția 68, 923-934. doi: 10.1111 / evo.12316
PubMed Abstract | CrossRef Full Text/Google Scholar
Edwards, S. V., Potter, S., Schmitt, C. J., Bragg, J. G. și Moritz, C. (2016). Reticularea, divergența și filogeografia–filogenetica continuumc. Proc. Natl. Acad. Sci. SUA 113, 8025-8032. doi: 10.1073 / pnas.1601066113
CrossRef Full Text/Google Scholar
Faith, D. P. (1992). Evaluarea conservării și diversitatea filogenetică. Biol. Contra. 61, 1–10. doi: 10.1016/0006-3207(92)91201-3
CrossRef Full Text | Google Scholar
Finston, T. L., Johnson, ms, Humphreys, W. F., Eberhard, S. M. și Halse, S. A. (2007). Speciația criptică în două genuri de amfipode subterane răspândite reflectă modelele istorice de drenaj într-un peisaj antic. Mol. Ecol. 16, 355–365. doi: 10.1111 / j. 1365-294X.2006.03123.x
PubMed Abstract | CrossRef Full Text/Google Scholar
Firestone, K. B., Elphinstone, ms, Sherwin, W. B. și Houlden, B. A. (1999). Structura populației filogeografice a tigrului quolls dasyurus maculatus (Dasyuridae: Marsupalia), un marsupial carnivor pe cale de dispariție. Mol. Ecol. 8, 1613–1625. doi: 10.1046 / j. 1365-294x.1999.00745.x
CrossRef Full Text/Google Scholar
fi Unq7er, C., Robinson, C. T. și Malard, F. (2018). Specii criptice ca fereastră în schimbarea de paradigmă a conceptului de specie. Mol. Ecol. 27, 613–635. doi: 10.1111 / mec.14486
PubMed Abstract | CrossRef Full Text/Google Scholar
Frankham, R. (2010). Provocări și oportunități ale abordărilor genetice ale conservării biologice. Biol. Conserv. 143,1919–1927. doi: 10.1016 / j. biocon.2010.05.011
CrossRef Full Text/Google Scholar
Frankham, R., Ballou, J. D., Dudash, M. R., Eldridge, MD, Fenster, C. B., Lacy, R. C. și colab. (2012). Implicațiile diferitelor concepte de specii pentru conservarea biodiversității. Biol. Conserv. 153, 25–31. doi: 10.1016 / j. biocon.2012.04.034
CrossRef Full Text/Google Scholar
Fraser, DJ, și Bernatchez, L. (2001). Conservarea evolutivă adaptivă: către un concept unificat pentru definirea unităților de conservare. Mol. Ecol. 10, 2741–2752. doi: 10.1046 / j.1365-294X.2001.t01-1-01411.x
PubMed Abstract | CrossRef Full Text/Google Scholar
Funk, W. C., Mckay, J. K., Hohenlohe, P. A., și Allendorf, F. W. (2012). Valorificarea genomica pentru delimitarea unităților de conservare. Tendințe Ecol. Evol. 27, 489–496. doi: 10.1016/j.copac.2012.05.012
PubMed Abstract | CrossRef Full Text/Google Scholar
Garnett, S. T. și Christidis, L. (2017). Anarhia taxonomică împiedică conservarea. Natură 546, 25-27. doi: 10.1038 / 546025a
PubMed Abstract | CrossRef Full Text/Google Scholar
Gippoliti, S., Cotterill, F. P. D., Zinner, D. și Groves, C. P. (2018). Impactul inerției taxonomice pentru Conservarea diversității ungulate africane: o privire de ansamblu. Biol. Rev. 93, 115-130. doi: 10.1111 / brv.12335
PubMed Abstract | CrossRef Full Text/Google Scholar
Grant, V. (1981). Speciația Plantelor. New York, NY: Columbia University Press.
PubMed Abstract/Google Scholar
Hamilton, C. W. și Reichard, S. H. (1992). Practica actuală în utilizarea subspecii, soiurilor și formei în clasificarea plantelor sălbatice. Taxonul 41, 485-498. doi: 10.2307/1222819
CrossRef Full Text/Google Scholar
Harvey, M. S., Rix, M. G., Framenau, V. W., Hamilton, Z. R., Johnson, ms, Teale, R. J. și colab. (2011). Protejarea celor nevinovați: studierea taxonilor endemici cu rază scurtă de acțiune îmbunătățește rezultatele conservării. Inversează. Syst. 25, 1–10. doi: 10.1071/IS11011
CrossRef Full Text/Google Scholar
Hei, J., Waples, R. S., Arnold, ml, Butlin, R. K. și Harrison, R. G. (2003). Înțelegerea și confruntarea incertitudinii speciilor în biologie și conservare. Tendințe Ecol. Evol. 18, 597–603. doi: 10.1016/j.copac.2003.08.014
CrossRef Full Text/Google Scholar
Hoban, S. M., Hauffe, H. C., Perez-Espona, S., Arntzen, J. W., Bertorelle, G., Bryja, J. și colab. (2013). Aducerea diversității genetice în prim-planul politicii și gestionării conservării. Conserv. Genet. Resour. 5, 593–598. doi: 10.1007 / s12686-013-9859-y
CrossRef Text Complet/Google Scholar
Isaac, N. J., Mallet, J. și Mace, G. M. (2004). Inflația taxonomică: influența sa asupra macroecologiei și conservării. Tendințe Ecol. Evol. 19, 464–469. doi: 10.1016/j.copac.2004.06.004
PubMed Abstract | CrossRef Full Text/Google Scholar
Isaac, N. J., Turvey, S. T., Collen, B., Waterman, C., și Baillie, J. E. (2007). Mamifere la margine: priorități de conservare bazate pe amenințare și filogenie. PLoS Unul 2: e296. doi: 10.1371/jurnal.pone.0000296
PubMed Abstract | CrossRef Full Text/Google Scholar
Jones, Mr, și bun, JM (2016). Captarea țintită în genomica evolutivă și ecologică. Mol. Ecol. 25, 185–202. doi: 10.1111 / mec.13304
PubMed Abstract | CrossRef Full Text/Google Scholar
Jorger, K. M., și Schr Unktifdl, M. (2013). Cum se descrie o specie criptică? Provocări practice ale taxonomiei moleculare. În față. Zool. 10:59. doi: 10.1186/1742-9994-10-59
PubMed Abstract | CrossRef Full Text/Google Scholar
Laikre, L., Allendorf, F., Aroner, L. C., Baker, S., Gregovich, D. P., Hansen, M. M. și colab. (2010). Neglijarea diversității genetice în punerea în aplicare a Convenției privind diversitatea biologică. Conserv. Biol. 24, 86–88. doi: 10.1111 / j. 1523-1739.2009. 01425.x
PubMed Abstract | CrossRef Full Text/Google Scholar
Laikre, L., Palm, S. și Ryman, N. (2005). Structura genetică a populației peștilor: implicații pentru gestionarea zonei costiere. Ambio 34,111-119. doi: 10.1579/0044-7447-34.2.111
PubMed Abstract | CrossRef Full Text/Google Scholar
laici, T., Laffan, S. W., Gonz Uniclez-Orozco, C. E., credință, D. P., Rosauer, D. F., Byrne, M. și colab. (2015). Filodiversitatea pentru a informa Politica de conservare: un exemplu Australian. Sci. Total Environ. 534, 131–143. doi: 10.1016 / j.scitotenv.2015.04.113
PubMed Abstract | CrossRef Full Text/Google Scholar
Leache, A. D. și Oaks, J. R. (2017). Utilitatea polimorfismului nucleotidic unic (SNP). date în filogenetică. Ann. Rev. Ecol. Syst. 48, 69–84. doi: 10.1146/annurev-ecolsys-110316-022645
CrossRef Full Text | Google Scholar
Llorens, T. M., Macdonald, B., McArthur, S., Coates, D. J., and Byrne, M. (2015). Disjunct, highly divergent genetic lineages within two rare Eremophila (Scrophulariaceae: Myoporeae). species in a biodiversity hotspot: implications for taxonomy and conservation. Biol. J. Linn. Soc. 177, 96–111. doi: 10.1111/boj.12228
CrossRef Full Text | Google Scholar
Love Stowell, S. M., Pinzone, C. A., and Martin, A. P. (2017). Depășirea barierelor în calea intervențiilor active pentru diversitatea genetică. Biodivers. Conserv. 26, 1753–1765. doi: 10.1007 / s10531-017-1330-z
CrossRef Text Complet/Google Scholar
Mace, G. M. (2004). Rolul taxonomiei în conservarea speciilor. Filos. Trans. R. Soc. B: Biol. Sci. 359, 711–719. doi: 10.1098 / rstb.2003.1454
PubMed Abstract | CrossRef Full Text/Google Scholar
Mallet, J. (2007). Specie hibridă. Natură 446, 279-283. doi: 10.1038 / nature05706
PubMed Abstract | CrossRef Full Text/Google Scholar
Mayr, E. (1942). Sistematica și Originea speciilor. New York, NY: Columbia University Press.
Google Scholar
Mee, J. A., Bernatchez, L., Reist, J. D., Rogers, SM și Taylor, EB (2015). Identificarea unităților desemnabile pentru prioritizarea conservării intraspecifice: o abordare ierarhică aplicată complexului speciilor de pești albi de lac (Coregonus spp.). Evol. Appl. 8, 423–441. doi: 10.1111 / eva.12247
PubMed Abstract | CrossRef Full Text | Google Scholar
Millar, M. A., and Byrne, M. (2013). Cryptic divergent lineages of Pultenaea pauciflora M.B. Scott (Fabaceae: Mirbelieae) exhibit different evolutionary history. Biol. J. Linn. Soc. 108, 871–881. doi: 10.1111/bij.12002
CrossRef Full Text | Google Scholar
Moritz, C. (1994). Defining evolutionarily significant units for conservation. Trends Ecol. Evol. 9, 373–375. doi: 10.1016/0169-5347(94)90057-4
CrossRef Full Text | PubMed Abstract
Moritz, C. (1999). Unități de conservare și translocații: strategii pentru conservarea proceselor evolutive. Hereditas 130, 217-228. doi: 10.1111 / j. 1601-5223.1999. 00217.x
CrossRef Text Complet/Google Scholar
Moritz, C. (2002). Strategii de protejare a diversității biologice și a proceselor evolutive care o susțin. Syst. Biol. 51, 238–254. doi: 10.1080 / 10635150252899752
PubMed Abstract | CrossRef Full Text/Google Scholar
Moritz, C. și Potter, S. (2013). Importanța unei perspective evolutive în planificarea politicii de conservare. Mol. Ecol. 22, 5969–5971. doi: 10.1111 / mec.12565
PubMed Abstract | CrossRef Full Text/Google Scholar
Moritz, C., Pratt, R. C., Bank, S., Bourke, G., Bragg, J. G., Doughty, P. și colab. (2018). Diversitatea descendenței criptice, divergența dimensiunii corpului și simpatia într-un complex de specii de șopârle australiene (Gehyra). Evoluția 27, 54-66. doi: 10.1111 / evo.13380
CrossRef Full Text/Google Scholar
Morrison, W. R., Lohr, J. L., Duchen, P., Wilches, R., Trujillo, D., Mair, M. și colab. (2009). Impactul schimbărilor taxonomice asupra conservării: Ucide, poate salva sau este doar irelevant? Biol. Contra. 142, 3201–3206. doi: 10.1016 / j. biocon.2009.07.019
CrossRef Full Text/Google Scholar
Murphy, N. P., Adams, M., Guzik, M. T. și Austin, A. D. (2013). Micro-endemism extraordinar în amfipodele de primăvară din deșertul Australian. Mol. Filogenet. Evol. 66, 645–653. doi: 10.1016 / j. ympev.2012.10.013
PubMed Abstract | CrossRef Full Text/Google Scholar
Murphy, N. P., King, R. A. și Delean, S. (2015). Specii, ESUs sau populații? Delimitarea și descrierea diversității criptice morfologic în amfipodele de primăvară din deșertul Australian. Inversează. Syst. 29, 457–467. doi: 10.1071/IS14036
CrossRef Full Text | Google Scholar
Nater, A., Mattle-Greminger, mp, Nurcahyo, A., Nowak, M. G., de Manual, M., Desai, T. și colab. (2017). Dovezi morfometrice, comportamentale și genomice pentru o nouă specie de urangutan. Curr. Biol Celular. 27, 3487–3498. doi: 10.1016/j.pui.2017.09.047
CrossRef Full Text/Google Scholar
Nicotra, A. B., Chong, C., Bragg, J. G., Ong, C. R., Aitken, N. C., Chuah, A., și colab. (2016). Populația și descompunerea filogenomică prin genotipare prin secvențiere în Pelargonium Australian. Mol. Ecol. 25, 2000–2014. doi: 10.1111 / mec.13584
PubMed Abstract | CrossRef Full Text/Google Scholar
Nistelberger, H., Byrne, M., Coates, D. și Roberts, JD (2014a). Structura filogeografică puternică într-un miliped indică vicarianța Pleistocenului între populațiile de pe formațiunile de fier în bandă din Australia semi-aridă. PLoS Unul 9: e0093038. doi: 10.1371/jurnal.pone.0093038
PubMed Abstract | CrossRef Full Text/Google Scholar
Nistelberger, H., Gibson, N., Macdonald, B., Tapper, S.-L., și Byrne, M. (2014b). Dovezi filogeografice pentru două refugii mesice într-un hotspot de biodiversitate. Ereditatea 113, 454-463. doi: 10.1038 / hdy.2014.46
PubMed Abstract | CrossRef Full Text | Google Scholar
Ogilvie, H. A., Heled, J., Xie, D. și Drummond, A. J. (2016). Performanța computațională și precizia statistică a beast și comparații cu alte metode. Syst. Biol. 65, 381–396. doi: 10.1093/sysbio/syv118
PubMed Abstract | CrossRef Full Text/Google Scholar
Oliver, P. M., Keogh, J. S. și Moritx, C. (2015). Noi abordări ale catalogării și înțelegerii diversității evolutive: o perspectivă din herpetologia australiană. Aust. J. Zool. 62, 417–430. doi: 10.1071/ZO14091
CrossRef Full Text/Google Scholar
Oliver, P. M., Laver, R. J., Demello Martins, F., Pratt, R. C., Hunjan, S. și Moritz, C. (2017). Un nou hotspot al endemismului vertebratelor și un refugiu evolutiv în Australia tropicală. Scafandru. Distrib. 23, 53–66. doi: 10.1111 / ddi.12506
CrossRef Full Text/Google Scholar
Oliver, P. M. și Lee, MSY (2010). Codurile botanice și zoologice împiedică cercetarea biodiversității prin descurajarea publicării de noi specii anonime. Taxonul 59, 1201-1205. doi: 10.2307/20773990
CrossRef Full Text/Google Scholar
Padial, J., Miralles, A., De La Riva, I. și Vences, M. (2010). Viitorul integrativ al taxonomiei. În față. Zool. 7:16. doi: 10.1186/1742-9994-7-16
PubMed Abstract | CrossRef Full Text/Google Scholar
Palsb Unktilll, P. J., B Unkticrub, M., și Allendorf, F. W. (2007). Identificarea unităților de management utilizând date genetice ale populației. Tendințe Ecol. Evol. 22, 11–16. doi: 10.1016/j.copac.2006.09.003
PubMed Abstract | CrossRef Full Text/Google Scholar
Patton, J. L. și Smith, M. F. (1994). Parafil, polifil și natura limitelor speciilor în goperii de buzunar (genul Thomomys). Syst. Biol. 43, 11–26. doi: 10.1093/sysbio / 43.1.11
CrossRef Full Text/Google Scholar
Pierson, J. C., Coates, D. J., Oostermeijer, J. G. B., Beissinger, Sr, Bragg, J. G., Sunnucks, P. și colab. (2016). Factori genetici în planurile de recuperare a speciilor amenințate pe trei continente. În față. Ecol. Environ. 14, 433–440. doi: 10.1002/taxă.1323
CrossRef Full Text/Google Scholar
Potter, S., Bragg, J. G., Peter, B. M., Bi, K. și Moritz, C. (2016). Filogenomica la sfaturi: deducând descendențele și istoria lor demografică într-o șopârlă tropicală, Carlia Amax. Mol. Ecol. 25, 1367–1380. doi: 10.1111 / mec.13546
PubMed Abstract | CrossRef Full Text/Google Scholar
Potter, S., Xue, A. T., Bragg, J. G., Rosauer, D. F., Roycroft, E. J. și Moritz, C. (2018). Schimbările climatice din Pleistocen determină diversificarea într-o savană tropicală. Mol. Ecol. 27, 520–532. doi: 10.1111 / mec.14441/full
PubMed Abstract | CrossRef Full Text/Google Scholar
Rabosky, D. L., Hutchinson, M. N., Donnellan, S. C., Talaba, A. L. și Lovette, I. J. (2014). Demontarea filogenetică a limitelor speciilor într-un grup larg răspândit de piele australiană (Scincidae: Ctenotus). Mol. Filogenet. Evol. 77, 71–82. doi: 10.1016 / j. ympev.2014.03.026
PubMed Abstract | CrossRef Full Text/Google Scholar
Ralls, K., Ballou, J. D., Dudash, Mr, Eldridge, MDB, Fenster, C. B., Lacy, RC, și colab. (2018). Solicită o schimbare de paradigmă în gestionarea genetică a populațiilor fragmentate. Conserv. Let. 11, 1–6. doi: 10.1111 / conl.124126
CrossRef Text Complet/Google Scholar
Rannala, B. (2015). Arta și știința delimitării speciilor. Curr. Zool. 61, 846–853. doi: 10.1093/czoolo/61.5.846
CrossRef Full Text/Google Scholar
Riedel, A., Sagata, K., Suhardjono, Y. R., T Inktoknzler, R., și Balke, M. (2013). Taxonomia Integrativă pe calea rapidă – către o mai mare durabilitate în cercetarea biodiversității. În față. Zool. 10:15. doi: 10.1186 / 1742-9994-10-15
PubMed Abstract | CrossRef Full Text/Google Scholar
Rieseberg, LH, și Willis, JH (2007). Speciația plantelor. Știință 317, 910-914. doi: 10.1126 / știință.1137729
PubMed Abstract | CrossRef Full Text/Google Scholar
Rosauer, D. F., Blom, MPK, Bourke, G., Catalano, S., Donnellan, S., Mulder, E. și colab. (2016). Filogeografie, puncte fierbinți și priorități de conservare: un exemplu din capătul superior al Australiei. Biol. Contra. 204, 83–93. doi: 10.1016 / j. biocon.2016.002
CrossRef Full Text/Google Scholar
Rosauer, D. F., Byrne, M., Blom, M. P. K., Coates, D. J., Donnellan, S., Doughty, P. și colab. (2018). Planificarea conservării în lumea reală pentru diversitatea evolutivă din Kimberley, Australia, evită taxonomia incertă Conservv. Let. 11: e12438. doi: 10.1111 / conl.12438
CrossRef Full Text/Google Scholar
Rosenblum, E. B., Sarver, B. A. J., Brown, J. W., Des Roches, S., Hardwick, K. M., Hether, T. D. și colab. (2012). Goldilocks întâlnește Santa Rosalia: un model de speciație efemer explică tiparele de diversificare pe scări de timp. Evol. Biol. 39, 255–261. doi: 10.1007 / s11692-012-9171-x
PubMed Abstract | CrossRef Full Text/Google Scholar
Roux, C., fra Inquxsse, C., Romiguier, J., Anciaux, Y., Galtier, N. și Bierne, N. (2016). Aruncând lumină asupra zonei gri a speciației de-a lungul unui continuum de divergență genomică. PLoS Biol. 14: e2000234. doi: 10.1371/jurnal.pbio.2000234
PubMed Abstract | CrossRef Full Text/Google Scholar
Ryder, O. A. (1986). Conservarea speciilor și sistematica – dilema subspecii. Tendințe Ecol. Evol. 1, 9–10. doi: 10.1016/0169-5347 (86)90059-5
CrossRef Full Text | Google Scholar
Sampson, J. F., Byrne, M., Yates, C. J., Gibson, N., Thavornkanlapachai, R., Stankowski, S. și colab. (2014). Imigrația genetică contemporană mediată de polen reflectă izolarea istorică a unui arbust rar într-un peisaj fragmentat. Ereditatea 112, 172-181. doi: 10.1038 / hdy.2013.89
PubMed Abstract | CrossRef Full Text | Google Scholar
Santamar INQUA, L., și m Inqqcndez, P. F. (2012). Evoluția politicii în domeniul biodiversității: lacunele actuale și nevoile viitoare. Evol. Appl. 5, 202–218. doi: 10.1111 / j. 1752-4571.2011.00229.x
PubMed Abstract | CrossRef Full Text/Google Scholar
Schlick-Steiner, B. C., Steiner, F. M., Seifert, B., Stauffer, C., Christian, E. și Crozier, R. H. (2010). Taxonomie Integrativă: o abordare multisource a explorării biodiversității. Anu. Rev. Entomol. 55, 421–438. doi: 10.1146 / annurev-ento-112408-085432
PubMed Abstract | CrossRef Full Text/Google Scholar
Seehausen, o, Takimoto, G., Roy, D. și Jokela, J. (2008). Inversarea speciației și dinamica biodiversității cu hibridizare în medii în schimbare. Mol Ecol. 17, 30–44. doi: 10.1111 / j. 1365-294X.2007.03529.x
PubMed Abstract | CrossRef Full Text/Google Scholar
Shepherd, K. A., Thiele, K. R., Sampson, J., Coates, DJ și Byrne, M. (2015). O specie rară, nouă de Atriplex (Chenopodiaceae) care cuprinde două populații distincte genetic, dar morfologic criptice în Australia de Vest aridă: implicații pentru taxonomie și conservare. Aust. Syst. Bot. 28, 234–245. doi: 10.1071/SB15029
CrossRef Text Complet/Google Scholar
Singhal, S. și Bi, K. (2017). Istoria curăță mizeria: impactul timpului în conducerea divergenței și introgresiei într-o zonă tropicală de sutură. Evoluție 71, 1888-1899. doi: 10.1111 / evo.13278
PubMed Abstract | CrossRef Full Text/Google Scholar
Singhal, S., Hoskin, C., Couper, P., Potter, S. și Moritz, C. (2018). Cadru pentru rezolvarea speciilor criptice: un studiu de caz din șopârlele tropicelor umede australiene. Syst. Biol. . doi: 10.1093/Sysbio | syy
PubMed Abstract/CrossRef Full Text/Google Scholar
Singhal, S., și Moritz, C. (2013). Izolarea reproductivă între liniile filogeografice scale cu divergență. Proc. Roy. Soc B 280: 20132246. doi: 10.1098 / rspb.2013.2246026
PubMed Abstract | CrossRef Full Text/Google Scholar
Steane, D. A., Potts, B. M., McLean, E., Collins, L., Prober, S. M., stoc, W. D., și colab. (2015). Scanările la nivel de genom dezvăluie structura criptică a populației într-un eucalipt adaptat la uscat. Arborele Genet. Genomul 11: 33. doi: 10.1007 / s11295-015-0864-z
CrossRef Full Text/Google Scholar
Struck, T. H., Feder, J. L., Bendiksby, M., Birkeland, S., Cerca, J., Gusarov, V. I. și colab. (2017). Găsirea proceselor evolutive ascunse în speciile criptice. Tendințe Ecol. Evol. 33, 153–163. doi: 10.1016/j.copac.2017.11.007
PubMed Abstract | CrossRef Full Text/Google Scholar
Sukumaran, J. și Knowles, L. L. (2017). Coalescența multispecifică delimitează structura nu speciile. Proc. Nat. Aca. Sci. SUA 114, 1607-1612. doi: 10.1073 / pnas.1607921114
PubMed Abstract | CrossRef Full Text/Google Scholar
Swarts, N. D., Clements, ma, Bower, C. C. și Miller, J. T. (2014). Definirea unităților de conservare într-un complex de orhidee similare morfologic, înșelătoare Sexual, foarte amenințate. Biol. Contra. 174, 55–64. doi: 10.1016 / j. biocon.2014.03.017
CrossRef Full Text/Google Scholar
Tautz, D., Arctander, P., Minelli, A., Thomas, R. H. și Vogler, A. P. (2003). O pledoarie pentru taxonomia ADN-ului. Tendințe Ecol. Evol. 18, 70–74. doi: 10.1016/S0169-5347(02)00041-1
CrossRef Text Complet/Google Scholar
Taylor, HR, Dussex, N., și van Heezik, Y. (2017). Reducerea decalajului geneticii de conservare prin identificarea barierelor în calea implementării pentru practicienii de conservare. Glob. Ecol. Conserv. 10, 231–242. doi: 10.1016 / j. gecco.2017.04.001
CrossRef Full Text/Google Scholar
Thomson, S. A., Pyle, R. L., Ahyong, S. T., Alonso-Zarazaga, M., Ammirati, J., Araya, J. F. și colab. (2018). Taxonomia bazată pe știință este necesară pentru conservarea globală. PLoS Biol. 16: e2005075. doi: 10.1371 / jurnal.pbio.2005075
PubMed Abstract | CrossRef Full Text/Google Scholar
Vieites, D. R., Wollenberg, K. C., Andreonec, F., K Inkthler, J., Glaw, F. și Vences, M. (2009). Subestimarea vastă a biodiversității Madagascarului evidențiată de un inventar integrativ de amfibieni. Proc. Natl. Acad. Sci. SUA 106, 8267-8272. doi: 10.1073 / pnas.0810821106
CrossRef text complet
Vogler, A. P. și de Salle, R. (1994). Diagnosticarea Unităților de gestionare a conservării. Contra. Biol. 8, 354–363. doi: 10.1046 / j. 1523-1739.1994. 08020354.x
CrossRef Full Text/Google Scholar
Waples, R. S. (1991). Somon de Pacific, Oncorhynchus spp., și definiția „speciilor” în temeiul Legii privind speciile pe cale de dispariție. Mar. Pește. Rev. 53, 11-22.
Google Scholar
Waples, R. S. (1995). „Unități semnificative din punct de vedere evolutiv și conservarea diversității biologice în temeiul Legii privind speciile pe cale de dispariție”, în evoluție și ecosistemul acvatic: definirea Unităților unice în conservarea populației, eds J. L. Nielsen (Bethesda, MD:American Fisheries Society), 8-27.
Google Scholar
Waples, R. S., Gustafson, R. G., Weitkamp, L. A., Myers, J. M., Jjonson, O. W., Busby, P. J. și colab. (2001). Caracterizarea diversității somonului din nord-vestul Pacificului. J. Pește Biol. 59, 1–41. doi: 10.1111 / j. 1095-8649.2001.tb01376.x
CrossRef Full Text/Google Scholar
Waples, R. S., Nammack, M., Cochrane, J. F. și Hutchings, J. A. (2013). O poveste despre două acte: practici de listare a speciilor pe cale de dispariție în Canada și Statele Unite. BioScience 63, 723-734. doi: 10.1093/știință biologică / 63.9.723
CrossRef Full Text/Google Scholar
Waples, R. S., Punt, A. E. și Cope, J. M. (2008). Integrarea datelor genetice în gestionarea resurselor marine: cum o putem face mai bine? Pește 9, 423-449. doi: 10.1111 / j. 1467-2979.2008. 00303.x
CrossRef Full Text/Google Scholar
Wege, J. A., Thiele, K. R., Shepherd, K. A., Butcher, R., Macfarlane, T. D. și Coates, D. J. (2015). Taxonomie strategică într-un peisaj biodivers: o abordare nouă pentru maximizarea rezultatelor conservării pentru flora rară și puțin cunoscută. Biodivers. Conserv. 24, 17–32. doi: 10.1007 / s10531-014-0785-4
CrossRef Text Complet/Google Scholar
Whitlock, R. (2014). Relațiile dintre diversitatea genetică adaptivă și neutră și structura și funcționarea ecologică: o meta-analiză. J. Ecol. 102, 857–872. doi: 10.1111 / 1365-2745.12240
PubMed Abstract | CrossRef Full Text/Google Scholar
Wiley, E. O. (1978). Conceptul de specie evolutivă a fost reconsiderat. Syst. Biol. 27, 17–26. doi: 10.2307/2412809
CrossRef Text Complet/Google Scholar
Wilson, E. O. (2017). Cercetarea biodiversității necesită mai multe cizme pe teren. Nat. Ecol. Evol. 1, 1590–1591. doi: 10.1038 / s41559-017-0360-y
PubMed Abstract | CrossRef Full Text/Google Scholar
Yang, Z. H. și Rannala, B. (2014). Delimitarea neguidată a speciilor folosind date de secvență ADN din mai multe loci. Mol. Biol. Evol. 31, 3125–3135. doi: 10.1093 / molbev / msu279
PubMed Abstract | CrossRef Full Text/Google Scholar
Yeates, D. K., Seago, A., Nelson, L., Cameron, S. L., Joseph, L. și Trueman, W. H. (2011). Taxonomie Integrativă sau taxonomie iterativă? Sys. Ent. 36, 209–217. doi: 10.1111/j.1365-3113.2010.00558.x
CrossRef Full Text | Google Scholar