i begynnelsen av XIX-tallet observert von Reckingausen at blodårene var dekket av et ark med celler. Dette encelle tykke laget består av endotelceller som strekker den indre overflaten av blod og lymfekar. Hos mennesker er det anslått at den totale endotellaget overflaten er ca 35 m2 og at det er rundt 1 til 1,6 103 endotelceller. Den totale lengden på arterier, vener og kapillærer er ca 90000 km (mer enn å reise to ganger rundt om i verden). Endotelceller er flate og forbundet mellom hverandre ved cellekryss. Deres hovedfunksjon er som mellommenn mellom blodet og det andre vevet, og bidrar til blodegenskaper, både i normalt og patologisk vev. De er også viktige for utveksling av immunceller mellom blod og vev.
1. Morfologi
Endotelceller er svært flattede celler, så mye at deres kjerne er den høyere strukturen til cellen, selv om kjernen også er flatt. Den cellulære formen er tilpasset kanalen endotelcellen er lining (Figur 1, 2 og 3). I de mer smale kapillærene kan endotelceller utvide sin cytoplasma hele blodkarets omkrets, slik at kanalen er en rekke endotelceller. I større kanaler, som arterier og vener, er det behov for mange endotelceller for å dekke kanalens totale omkrets.
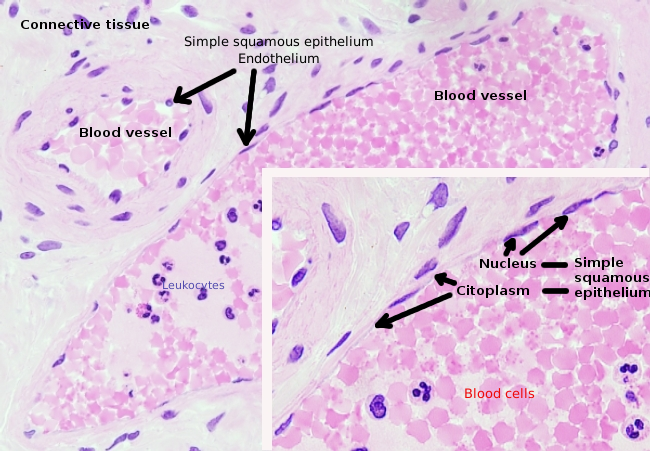
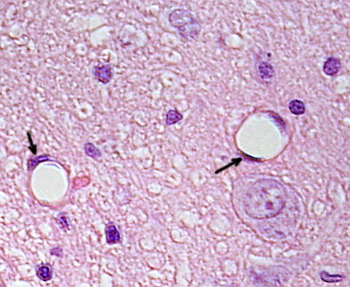
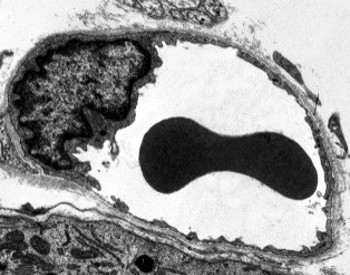
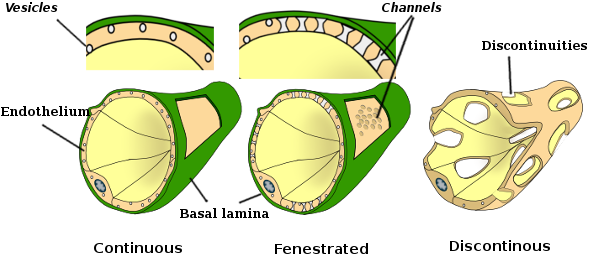
Perinukleær cytoplasma inneholder de fleste mitokondrier og andre organeller, som Golgi-apparater, mens perifericytoplasma er tynnere og har få organeller, selv om endoplasmatisk retikulum kan finnes nær plasmamembranen. Endotelceller har mange vesikler, de fleste er endocytiske vesikler. I noen regioner, som lever, nyrebark og endokrine kjertler, viser endotelceller av kapillærer porer eller små passasjer, kjent som fenestratrioner. De er veier på ca 60 til 70 nm i diameter, med en smal passasje på ca 5 nm, som direkte kommuniserer blod og lymf med de andre vevene. Kapillærer som har disse typer endotelceller er kjent som fenestrerte kapillærer. Porene fordeles i grupper og dens tetthet avhenger av typen endotel.
Endotelceller separeres fra det omgivende vevet av et lag av ekstracellulær matrise kalt basal lamina. Endotelcelle bidrar til dette laget med proteinene laminin, fibronektin og kollagen TYPE II, IV OG V.
Kilde og proliferasjon
Vaskulogenese er dannelsen av nye endotelceller i vev uten eksisterende blodkar. Det skjer i embryoet. Hos voksne, og også i embryoer, genereres nye blodkar, og derfor nye endotelceller, fra allerede tilstedeværende blodkar. Denne prosessen er kjent som angiogenese. Nye endotelceller produseres ved forgrening eller deling av blod og lymfekar. Angiogenese er vanlig i voksende vev eller de som er under tung remodeling, som livmor hos pattedyrhunner, eller i patologier som svulster, betennelser og sår.
Vaskulogenese
Første endotelceller av pattedyr genereres etter gastrulering under embryoutviklingen. De skiller seg fra celler som kalles angioblaster, danner grupper og blir organisert i korte blodkar. Denne prosessen skjer først i vitelin sac av embryoet. Disse innledende blodkar vokse og koble mellom hverandre for å danne et nettverk. Senere rekrutterer de fibroblaster og muskelceller. Dette første nettverket gjennomgår en kontinuerlig ombyggingsprosess i de neste utviklingsstadiene. I embryoer er alle endotelceller i utgangspunktet like, men de blir senere differensiert i vene, arterie, kapillær og lymfatisk karendotel. Flere molekyler som FGF2, BMP4, IHH og VEGF induserer separerte differensieringsveier. Notch inducer synes å være svært viktig fordi det fører til arterie endotel, mens dens hemming fører til veneendotel. Lymfatisk endotel er dannet fra kardinal veneendotelet og trenger uttrykk FOR COUPF-II OG SOX18. I tillegg er endotelet differensiert tilsvarende til orgelet det ligger i. For eksempel danner endotelet et tett forseglet lag i hjernen for å danne blod-hjernebarrieren, mens det er fenestrert av løst organisert i leveren for å favorisere utveksling av molekyler med blodet.som endotelceller blir organisert i arterie-og venekanaler, gir noen endotelceller opphav til hematopoietiske celler ved en differensieringsprosess kjent som endotelial-hematopoietisk overgang (EHT). Denne prosessen skjer også i noen voksne vev som under dannelsen av placenta og andre vev under svangerskapet.
Angiogenese
Angiogenese er dannelsen av nye blodkar og endotel ved å forgrene eller dele eksisterende blodkar. Hos voksne er proliferasjonshastigheten av endotelceller svært lav. For eksempel, i pattedyr, kan en endotelcelle dele seg en gang per måned eller kan vente flere år. Det betyr at angiogenese er sjelden i vev under normal fysiologi, unntatt kvinnelige reproduktive organer. Det kan imidlertid aktiveres i patologiske prosesser som vevskader og svulster. Noen stoffer induserer angiogenese, som endotelvekstfaktor (EGF), sur EGF, fibroblastvekstfaktor (FGF), transformerende vekstfaktor (TGF) og prostaglandiner. Angiogenese begynner når angiogene stoffer frigjøres fra omgivende vev. Endotelcellen i et nær blodkar, vanligvis fra en venule, begynner å migrere til den angiogene signalkilden, dra de andre endotelceller, som begynner å proliferere og danne et nytt fartøy. Disse endotelceller mister forbigående sine tette forbindelser som resulterer i avdekking av basalmembranen, som raskt fordøyes av enzymer frigjort fra endotelceller. Noen endotelceller går gjennom den fordøyede basalmembranen mens spredning fortsetter. All denne prosessen muliggjør forlengelse og forgrening av det nye blodkaret til vaskulogenesesignalet forsvinner. Spissene i blodårene smelter sammen med andre blodkar for å danne lukkede kretser og tillate fluss av blod uten døde endinger.
Regioner med endotelcelleforfedre har blitt funnet i dorsal aorta og endokardium. Mesenkymceller kan differensieres fra endokardiumendotelceller og danne tricuspidventilen og noen fibroblaster i hjertet, men ikke kardiomiocytter. Endotelceller er så fornuftige for eksterne signaler at det å holde endotelintegriteten er en aktiv prosess, som formidles av andre signaler som FGF. Den aktive prosessen er ikke bare for integritet, men for å holde den cellulære fenotypen også. Når endotelceller ikke mottar riktige signaler, dør de av apoptose eller blir mesenkymale celler som kan syntetisere rikelig ekstracellulær matrise. Overgangen fra endotel til mesenkymal celle ligger bak noen sykdommer som arteriosklerose og fibrøs miokarditt.
Funksjon
tanken om at endotelet bare er et passivt foringslag av lymfatiske og blodkar må endres. Endotelfunksjoner er varierte og essensielle for organismen. Faktisk er det en fysisk struktur av kardiovaskulære og lymfatiske kanaler, men det regulerer også utvekslingen av molekyler mellom fartøyets lumen og det omkringliggende vevet, og det påvirker blodets fysiologiske egenskaper. Foruten en fysisk barriere utvikler endotelceller sekretoriske, metabolske og immunfunksjoner. De endrer sin fysiologiske oppførsel påvirket av molekyler som vekstfaktorer, koagulanter og antikoagulantia, lipoproteiner med lav tetthet, nitrogenoksid, serotonin, enkefalin og mange andre. Endotelceller har reseptorer for alle disse stoffene.
Barriere
Endotelceller danner et lag som vanligvis virker som en barriere mellom blodet og det omkringliggende vevet. Kohesjonen mellom endotelceller oppfylles av cellekryss, for eksempel tette veikryss og vedheftende veikryss. Gapkryss har også blitt observert, selv om deres hovedfunksjon er kommunikasjonen mellom tilstøtende celler. Endotelceller kan modulere disse adhesjonene og endre permeabiliteten til barrieren, som kan påvirke ikke bare molekyler, men også celler som krysser endotelet. Men i noen organer som lever er endotelceller mye mer løst pakket og gir mye ledig plass, slik at endotelet nesten ikke kan betraktes som en barriere.
Transmisjonselektronmikroskopibilder viser mange vesikler i cytoplasma av endotelceller. De antas å være involvert i intracellulær transport mellom apikalen (mot blodet) og basolaterale membraner (mot basallamina). Denne typen transport er kjent som transcytose. Det er interessant at transcytose vesikler er mer rikelig i endotelceller i kapillærene enn i de av større blodkar. Det indikerer at endotelceller i kapillærene har en mer intens utveksling av molekyler mellom blod og vev, mens de i stor diameter er hovedsakelig involvert i å gjennomføre blodet. Noen endotelceller, kjent som fenestrert, har porer eller svært små kanaler som direkte forbinder blodet med det omkringliggende vevet, slik at noen småstørrelsesmolekyler kan krysse endotelet uten å komme inn i cytoplasma av noen endotelceller (Figur 4). Til slutt er det organer med svært permeabelt endotel. I leveren er sinusoider blodkar hvor endotelceller gir ledig plass mellom hverandre, og en funksjon som barriere eksisterer knapt.
Mange celletyper reiser i blodet mot målorganene i kroppen. Der krysser de endotelet i blodkarene, vanligvis på nivået av post-kapillære årer. Det betyr at endotelceller må modifisere cellekryssene for å la celler gå gjennom endotellaget. Leukocytter går ut av blodkaret ved å gjenkjenne og forankre til bestemte molekyler av den apikale membranen i endotelceller. Selektiner, integriner og immunglobuliner er ansvarlige for gjenkjenning og adhesjon av leukocytter til endotellaget. Selectins begynner forankring av leukocytter, som ruller over endoteloverflaten. Denne første adhesjonen er svak og reversibel.
mange leukocytter går ut av blodet under inflammatoriske prosesser for å bevege seg til de berørte vevene. Kjemokiner er leukocytt-attraktive molekyler som frigjøres av skadede vev og knyttes til glycocalix av endotelceller. Leukocytter som ruller over endoteloverflaten med kjemokiner aktiveres, cellecelleadhesjon blir sterkere, og leukocytt forblir på plass. Aktivering av leukocytten fører til aktivering av integrinene, som gjenkjenner immunoglobuliner av endotelcellen. Disse immunoglobuliner uttrykkes i endotelcellemembranene etter aktivering av cellen av kjemokiner. Celle-celle adhesjon øker kalsiumkonsentrasjonen i endotelceller og fører til celle veikryss disorganisering og cytoplasma tilbaketrekning. På denne måten kan leukocytter bevege seg til grensen til endotelceller og krysse endotelet. Adhesjonsmolekyler er også involvert i denne bevegelsen av leukocytter.
blodegenskaper
endotelfunksjonene er mer komplekse enn bare å kontrollere molekylene og cellene som går over endotelet. Endotelceller er også involvert i blodtrykk, koagulasjon og noen andre blodegenskaper. Primitivt sirkulasjonssystem antas å dukke opp for 600 millioner år siden i hvirvelløse dyr, men det manglet endotel. Endotelceller dukket opp for 100 millioner år siden, og ga blod med en mer laminær flux (ikke turbulent) og dermed en mer effektiv gassutveksling.
Endotelceller modulerer blodtrykket ved å frigjøre stoffer som virker på glatt muskel i blodårene. De frigjør nitrogenoksid (NO) og prostacyklin, som slapper av vaskulær glatt muskel. De frigjør også endotelin og blodplateaktivatorfaktoren, som begge reduserer blodkarets diameter. Nitrogenoksid frigjøres konstitusjonelt og gir en riktig muskelton, hemmer blodplateaggregering og leukocyttadhesjon. Endotelin er en sterk vasokonstriktor. Hvordan endotelceller bestemmer hvilket molekyl som skal frigjøres, er ikke klart ennå, men mekanorreceptorer i deres apikale membraner som er i stand til å føle blodets fluxegenskaper, kan være plausible. under normale forhold frigjør endotelceller molekyler i blodet som bidrar til å opprettholde en riktig fluiditet. De arbeider på to nivåer: fluiditet (antikoagulantia) og forebygging av blodplateaggregering (antitrombotikk). Proteiner C og S er viktige molekyler som påvirker blodfluiditeten. C-protein, som danner et kompleks Med s-protein, inaktiverer koagulasjonsfaktorer VIIIa Og Va. S protein syntetiseres av endotelceller. Videre inneholder endotelet glycocalix en glykosaminoglykan lik heparin som er i stand til å inaktivere trombin. Relatert til deres antikoagulerende funksjon frigjør endotelceller nitrogenoksid og prostacyklin, som både øker den sykliske AMP i blodplater og gjør vanskeligere blodplateaggregasjon. Disse to molekylene frigjøres kontinuerlig i blodet. Endotelceller har ektonukleaser i deres apikale membraner, som fjerner ATP og ADP, begge sterke promotorer av blodplateaggregering. I tillegg frigjør endotelceller et aktivatormolekyl som forvandler plaminogen til plasmin, noe som favoriserer fjerning av trombi.
Alle disse molekylære veiene kan endres når endotelceller mottar noen signaler eller vev er skadet, noe som fører til blodkoagulasjon og blodplateaggregering. Under disse omstendighetene blir endotelceller aktive deltakere i koagulasjon og trombose.
Immunforsvar
Endotelceller spiller en viktig rolle i immunforsvaret og er involvert i to mekanismer: antigenerpresentasjon Til T-lymfocytter og rekruttering av immunceller. Sammen med makrofager kan endotelceller presentere antigener til T-lymfocytter fordi DE konstitutivt uttrykker MHC-i (major histocompatibility complex) og kan induseres til å uttrykke MHC-II, begge nødvendige for antigenpresentasjon. Endotelceller er i stand til å aktivere immunforsvar, men ikke nye t-lymfocytter. Det er toveis aktivering mellom endotelcelle Og T-lymfocytter, slik at endotelcellefrigjøringsmolekyler for å tiltrekke inflammatoriske celler og uttrykke adhesjonsmolekyler for forankring av blodleukocytter.
Bibliografi
Cines BD, Pollak ES, Buck CA, Loscalzo J, Zimmerman GA, McEver RP, Pober JS, Wick TM, Konkle BA, Schwartz BS, Barnathan ES, McCrae KR, Klem BA, Schmidt A-M, Stern dm 1998. Endotelceller i fysiologi og i patofysiologiav vaskulære lidelser. Tidsskrift for den norske legeforening. 91:3527-3561![]()
Fajardo LF. 1988. Kompleksiteten av endotelceller. American journal of clinical pathology (engelsk). 92:241-250.
Michiels C. 2003. Endotelcellefunksjoner. Tidsskrift for rettsvitenskap. 196:430-443![]()