All’inizio del XIX secolo, von Reckingausen osservò che i vasi sanguigni erano coperti da un foglio di cellule. Questo strato spesso a una cellula è costituito da cellule endoteliali che rivestono la superficie interna dei vasi sanguigni e linfatici. Negli esseri umani, si stima che la superficie totale dello strato endoteliale sia di circa 35 m2 e che ci siano da 1 a 1,6 103 cellule endoteliali. La lunghezza totale di arterie, vene e capillari è di circa 90000 km (più di viaggiare due volte in tutto il mondo). Le cellule endoteliali sono piatte e collegate tra loro da giunzioni cellulari. La loro funzione principale è come intermediari tra il sangue e gli altri tessuti e contribuiscono alle proprietà del sangue, sia nei tessuti normali che patologici. Sono anche importanti per lo scambio di cellule immunitarie tra sangue e tessuti.
1. Morfologia
Le cellule endoteliali sono cellule molto appiattite, tanto che il loro nucleo è la struttura più alta della cellula, anche se anche il nucleo è appiattito. La forma cellulare è adattata al condotto che la cellula endoteliale sta rivestendo (Figura 1, 2 e 3). Nei capillari più stretti, le cellule endoteliali possono estendere il loro citoplasma su tutto il perimetro del vaso sanguigno, in modo che il condotto sia una fila di cellule endoteliali. Nei condotti più grandi, come arterie e vene, sono necessarie molte cellule endoteliali per coprire il perimetro totale del condotto.
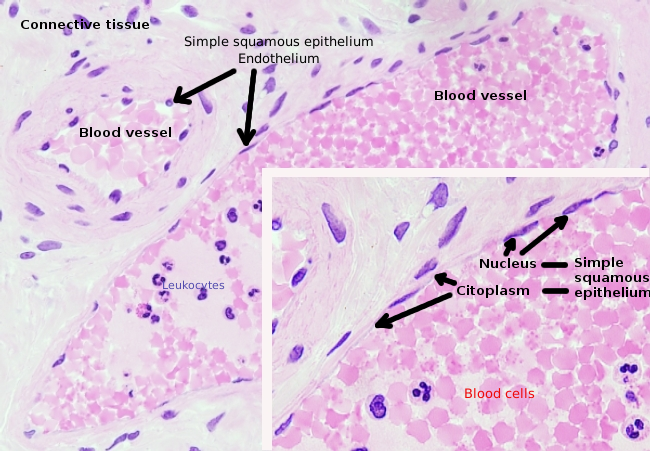
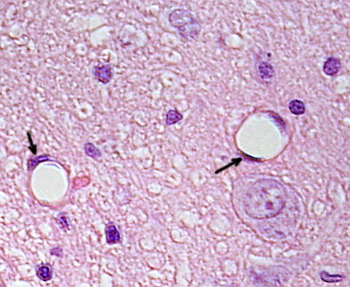
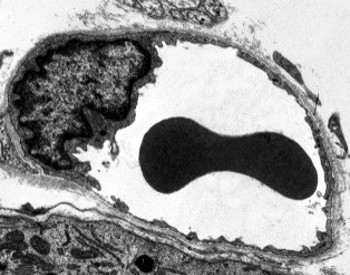
Il citoplasma perinucleare contiene la maggior parte dei mitocondri e altri organelli, come l’apparato di Golgi, mentre il citoplasma periferico è più sottile e ha pochi organelli, sebbene il reticolo endoplasmatico possa essere trovato vicino alla membrana plasmatica. Le cellule endoteliali hanno molte vescicole, la maggior parte di esse sono vescicole endocitiche.
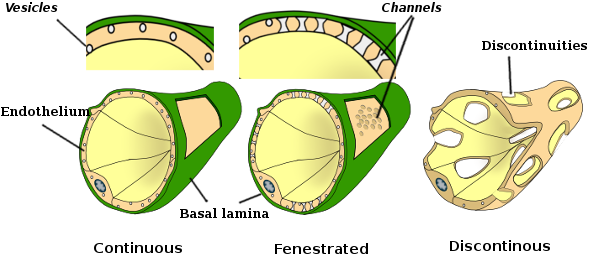
In alcune regioni, come il fegato, la corteccia renale e le ghiandole endocrine, le cellule endoteliali dei capillari mostrano pori o piccoli passaggi, noti come fenestratrioni. Sono percorsi di circa 60-70 nm di diametro, con uno stretto passaggio di circa 5 nm, che comunicano direttamente il sangue e la linfa con gli altri tessuti. I capillari che hanno questi tipi di cellule endoteliali sono noti come capillari fenestrati. I pori sono distribuiti in gruppi e la loro densità dipende dal tipo di endotelio.
Le cellule endoteliali sono separate dal tessuto surronding da uno strato di matrice extracellulare chiamato lamina basale. Le cellule endoteliali contribuiscono a questo strato con le proteine laminina, fibronectina e collagene di tipo II, IV e V.
Fonte e proliferazione
La vasculogenesi è la formazione di nuove cellule endoteliali nei tessuti senza vasi sanguigni preesistenti. Succede nell’embrione. Negli adulti, e anche negli embrioni, nuovi vasi sanguigni, e quindi nuove cellule endoteliali, vengono generati da vasi sanguigni già presenti. Questo processo è noto come angiogenesi. Le nuove cellule endoteliali sono prodotte ramificando o dividendo i vasi sanguigni e linfatici. L’angiogenesi è comune nei tessuti in crescita o in quelli sottoposti a rimodellamento pesante, come l’utero nelle femmine di mammifero, o in patologie come tumori, infiammazioni e ferite.
Vasculogenesi
Le prime cellule endoteliali dei mammiferi vengono generate dopo gastrulazione durante lo sviluppo dell’embrione. Si differenziano dalle cellule chiamate angioblasti, formano gruppi e si organizzano in brevi vasi sanguigni. Questo processo avviene per la prima volta nel sacco vitelin dell’embrione. Questi vasi sanguigni iniziali crescono e si connettono tra loro per formare una rete. Più tardi, reclutano fibroblasti e cellule muscolari. Questa rete iniziale subisce un continuo processo di rimodellamento durante le successive fasi di sviluppo. Negli embrioni, tutte le cellule endoteliali sono inizialmente simili, ma vengono successivamente differenziate in endotelio venoso, arterioso, capillare e linfatico. Diverse molecole come FGF2, BMP4, IHH e VEGF inducono percorsi di differenziazione separati. L’induttore di tacca sembra essere molto importante perché conduce all’endotelio dell’arteria, mentre la sua inibizione conduce all’endotelio della vena. L’endotelio linfatico è formato dall’endotelio della vena cardinale e ha bisogno dell’espressione di COUPF-II e SOX18. Inoltre, l’endotelio è differenziato di conseguenza all’organo in cui si trova. Ad esempio, l’endotelio forma uno strato ermeticamente sigillato nel cervello per formare la barriera emato-encefalica, mentre è fenestrato di vagamente organizzato nel fegato per favorire lo scambio di molecole con il sangue.
Poiché le cellule endoteliali si organizzano nei dotti arteriosi e venosi, alcune cellule endoteliali danno origine a cellule ematopoietiche mediante un processo di differenziazione noto come transizione endoteliale-ematopoietica (EHT). Questo processo avviene anche in alcuni tessuti adulti come durante la formazione di placenta e altri tessuti durante la gravidanza.
Angiogenesi
L’angiogenesi è la formazione di nuovi vasi sanguigni ed endotelio ramificando o dividendo i vasi sanguigni preesistenti. Negli adulti, il tasso di proliferazione delle cellule endoteliali è molto basso. Ad esempio, nei mammiferi, una cellula endoteliale può dividersi una volta al mese o può attendere diversi anni. Significa che l’angiogenesi è rara nei tessuti in condizioni normali di fisiologia, ad eccezione degli organi riproduttivi femminili. Tuttavia, può essere attivato in processi patologici come danni ai tessuti e tumori. Alcune sostanze inducono l’angiogenesi, come il fattore di crescita endoteliale (EGF), l’EGF acido, il fattore di crescita dei fibroblasti (FGF), il fattore di crescita trasformante (TGF) e le prostaglandine. L’angiogenesi inizia quando le sostanze angiogeniche vengono rilasciate dai tessuti circostanti. La cellula endoteliale di un vaso sanguigno vicino, di solito da una venula, inizia a migrare verso la sorgente del segnale angiogenico, trascinando le altre cellule endoteliali, che inizia a proliferare e formare un nuovo vaso. Queste cellule endoteliali perdono transitoriamente le loro connessioni strette con conseguente scoperta della membrana basale, che viene rapidamente digerita dagli enzimi rilasciati dalle cellule endoteliali. Alcune cellule endoteliali passano attraverso la membrana basale digerita mentre la proliferazione continua. Tutto questo processo rende possibile l’allungamento e la ramificazione del nuovo vaso sanguigno fino alla scomparsa del segnale di vasculogenesi. Le punte dei vasi sanguigni si fondono con altri vasi sanguigni per formare circuiti chiusi e consentire il flusso di sangue senza terminazioni morte.
Regioni con progenitori delle cellule endoteliali sono state trovate nell’aorta dorsale e nell’endocardio. Le cellule mesenchimali possono essere differenziate dalle cellule endoteliali dell’endocardio e formare la valvola tricuspide e alcuni fibroblasti del cuore, ma non i cardiomiociti. Le cellule endoteliali sono così sensibili ai segnali esterni che mantenere l’integrità dell’endotelio è un processo attivo, mediato da altri segnali come FGF. Il processo attivo non è solo per l’integrità, ma per mantenere il fenotipo cellulare pure. Quando le cellule endoteliali non ricevono segnali adeguati, muoiono per apoptosi o diventano cellule mesenchimali che possono sintetizzare un’abbondante matrice extracellulare. La transizione dalla cellula endoteliale a quella mesenchimale è alla base di alcune malattie come l’arteriosclerosi e la miocardite fibrosa.
Funzione
L’idea che l’endotelio sia solo un rivestimento passivo di vasi linfatici e sanguigni deve essere cambiata. Le funzioni dell’endotelio sono varie ed essenziali per l’organismo. Infatti, è una struttura fisica dei dotti cardiovascolari e linfatici, ma regola anche lo scambio di molecole tra il lume del vaso e i tessuti circostanti e influenza le proprietà fisiologiche del sangue. Oltre a una barriera fisica, le cellule endoteliali sviluppano funzioni secretorie, metaboliche e immunitarie. Cambiano il loro comportamento fisiologico influenzato da molecole come fattori di crescita, coagulanti e anticoagulanti, lipoproteine a bassa densità, ossido nitrico, serotonina, encefalina e molti altri. Le cellule endoteliali hanno recettori per tutte queste sostanze.
Barriera
Le cellule endoteliali formano uno strato che di solito funge da barriera tra il sangue e i tessuti circostanti. La coesione tra le cellule endoteliali è soddisfatta da giunzioni cellulari, come giunzioni strette e giunzioni aderenti. Sono state osservate anche giunzioni di gap, sebbene la loro funzione principale sia la comunicazione tra cellule adiacenti. Le cellule endoteliali possono modulare queste aderenze e modificare la permeabilità della barriera, che può influenzare non solo le molecole ma anche le cellule che attraversano l’endotelio. Tuttavia, in alcuni organi come il fegato, le cellule endoteliali sono molto più vagamente confezionate e lasciano molto spazio libero in modo che l’endotelio possa difficilmente essere considerato una barriera.
Le immagini di microscopia elettronica a trasmissione mostrano molte vescicole nel citoplasma delle cellule endoteliali. Si pensa che siano coinvolti nel trasporto intracellulare tra le membrane apicali (rivolte verso il sangue) e basolaterali (rivolte verso la lamina basale). Questo tipo di trasporto è noto come transcitosi. È interessante notare che le vescicole transcitosi sono più abbondanti nelle cellule endoteliali dei capillari che in quelle dei vasi sanguigni più grandi. Indica che le cellule endoteliali nei capillari hanno uno scambio più intenso di molecole tra sangue e tessuti, mentre nei vasi di grande diametro sono principalmente coinvolti nella conduzione del sangue. Alcune cellule endoteliali, note come fenestrate, hanno pori o condotti molto piccoli che collegano direttamente il sangue con i tessuti circostanti, consentendo ad alcune molecole di piccole dimensioni di attraversare l’endotelio senza entrare nel citoplasma di alcuna cellula endoteliale (Figura 4). Infine, ci sono organi con endotelio molto permeabile. Nel fegato, i sinusoidi sono vasi sanguigni in cui le cellule endoteliali lasciano spazio libero tra loro e una funzione come barriera difficilmente esiste.
Molti tipi di cellule viaggiano nel sangue verso i loro organi bersaglio nel corpo. Lì, attraversano l’endotelio dei vasi sanguigni, comunemente a livello delle vene post-capillari. Significa che le cellule endoteliali devono modificare le giunzioni cellulari per far passare le cellule attraverso lo strato di endotelio. I leucociti escono dal vaso sanguigno riconoscendo e ancorando a specifiche molecole della membrana apicale delle cellule endoteliali. Selectine, integrine e immunoglobuline sono responsabili del riconoscimento e dell’adesione dei leucociti allo strato endoteliale. Le selectine iniziano l’ancoraggio dei leucociti, che rotolano sulla superficie endoteliale. Questa adesione iniziale è debole e reversibile.
Molti leucociti escono dal sangue durante i processi infiammatori per spostarsi nei tessuti interessati. Le chemochine sono molecole attraenti per i leucociti che vengono rilasciate dai tessuti danneggiati e collegate al glicocalice delle cellule endoteliali. I leucociti che rotolano sulla superficie endoteliale con chemochine vengono attivati, l’adesione cellulare-cellulare diventa più forte e i leucociti rimangono nel posto. L’attivazione del leucocita porta all’attivazione delle sue integrine, che riconoscono le immunoglobuline della cellula endoteliale. Queste immunoglobuline sono espresse nelle membrane cellulari endoteliali dopo l’attivazione della cellula da parte delle chemochine. L’adesione cellula-cellula aumenta la concentrazione di calcio nelle cellule endoteliali e porta alla disorganizzazione delle giunzioni cellulari e alla retrazione del citoplasma. In questo modo, i leucociti possono spostarsi verso il bordo delle cellule endoteliali e attraversare l’endotelio. Le molecole di adesione sono anche coinvolte in questo movimento dei leucociti.
Proprietà del sangue
Le funzioni endoteliali sono più complesse del semplice controllo delle molecole e delle cellule che attraversano l’endotelio. Le cellule endoteliali sono anche coinvolte nella pressione sanguigna, nella coagulazione e in alcune altre proprietà del sangue.
Si pensa che il sistema circolatorio primitivo sia emerso 600 milioni di anni fa negli invertebrati, ma mancava di endotelio. Le cellule endoteliali apparvero 100 milioni di anni fa fornendo al sangue un flusso più laminare (non turbolento) e quindi uno scambio di gas più efficiente.
Le cellule endoteliali modulano la pressione sanguigna rilasciando sostanze che agiscono sulla muscolatura liscia dei vasi sanguigni. Rilasciano ossido nitrico (NO) e prostaciclina, che rilassano la muscolatura liscia vascolare. Rilasciano anche l’endotelina e il fattore attivatore piastrinico, diminuendo entrambi il diametro dei vasi sanguigni. L’ossido nitrico è costitutivamente rilasciato e fornisce un tono muscolare adeguato, inibisce l’aggregazione piastrinica e l’adesione dei leucociti. L’endotelina è un forte vasocostrittore. Come le cellule endoteliali decidano quale molecola deve essere rilasciata non è ancora chiaro, ma i meccanorecettori nelle loro membrane apicali che sono in grado di sentire le proprietà del flusso sanguigno possono essere plausibili.
In condizioni normali, le cellule endoteliali rilasciano molecole nel sangue che aiutano a mantenere una corretta fluidità. Funzionano a due livelli: fluidità (anticoagulanti) e prevenzione dell’aggregazione piastrinica (antitrombotici). Le proteine C e S sono molecole importanti che influenzano la fluidità del sangue. La proteina C, formando un complesso con la proteina S, inattiva i fattori di coagulazione VIIIa e Va. La proteina S è sintetizzata dalle cellule endoteliali. Inoltre, il glicocalice endoteliale contiene un glicosaminoglicano simile all’eparina che è in grado di inattivare la trombina.
In relazione alla loro funzione anticoagulante, le cellule endoteliali rilasciano ossido nitrico e prostaciclina, aumentando l’AMP ciclico nelle piastrine e rendendo più difficile l’aggregazione piastrinica. Queste due molecole vengono continuamente rilasciate nel sangue. Le cellule endoteliali hanno ectonucleasi nelle loro membrane apicali, che rimuovono ATP e ADP, entrambi forti promotori dell’aggregazione piastrinica. Inoltre, le cellule endoteliali rilasciano una molecola attivatrice che trasforma il plaminogeno in plasmina, che favorisce la rimozione dei trombi.
Tutte queste vie molecolari possono cambiare quando le cellule endoteliali ricevono alcuni segnali o i tessuti sono danneggiati, che portano alla coagulazione del sangue e all’aggregazione piastrinica. In queste circostanze, le cellule endoteliali diventano quindi partecipanti attivi della coagulazione e della trombosi.
Difesa immunitaria
Le cellule endoteliali svolgono un ruolo importante nella difesa immunitaria e sono coinvolte in due meccanismi: la presentazione degli antigeni ai linfociti T e il reclutamento delle cellule immunitarie. Insieme ai macrofagi, le cellule endoteliali possono presentare antigeni ai linfociti T perché esprimono costitutivamente MHC-I (complesso maggiore di istocompatibilità) e possono essere indotte ad esprimere MHC-II, entrambi necessari per la presentazione dell’antigene. Le cellule endoteliali sono in grado di attivare la memoria immunitaria, ma non nuovi linfociti T. C’è attivazione bidirezionale fra la cellula endoteliale ed i linfociti T, di modo che le molecole del rilascio delle cellule endoteliali per attirare le cellule infiammatorie ed esprimono le molecole di adesione per l’ancoraggio dei leucociti del sangue.
Bibliografia
Cines BD, Pollak ES, Buck CA, Loscalzo J, Zimmerman GA, McEver RP, Pober JS, Wick TM, Konkle BA, Schwartz BS, Barnathan ES, McCrae KR, Hug BA, Schmidt A-M, Stern DM 1998. Cellule endoteliali nella fisiologia e nella fisiopatologia dei disturbi vascolari. Il giornale della società americana di ematologia. 91:3527-3561 ![]()
Fajardo LF. 1988. La complessità delle cellule endoteliali. American journal of clinical pathology. 92:241-250.
Michiels C. 2003. Funzioni delle cellule endoteliali. Journal of cellular physiology. 196:430-443 ![]()