Johdanto
kansainvälisessä suojelupolitiikassa tunnustetaan biologinen monimuotoisuus kolmella tasolla, ekosysteemi, lajit ja geneettinen monimuotoisuus, ja että hoidon tavoitteena olisi oltava kaikkien kolmen säilyttäminen (biologista monimuotoisuutta koskeva yleissopimus, 2007). Tämä näkyy selvästi vuonna 2010 hyväksytyssä biologista monimuotoisuutta koskevassa yleissopimuksessa Aichi Biodiversity objectives (https://www.cbd.int/sp/targets/), jossa tavoitteissa ja tavoitteissa viitataan erityisesti ekosysteemien ja lajien lisäksi geneettiseen monimuotoisuuteen. Nykyiset lähestymistavat biologisen monimuotoisuuden säilyttämiseen perustuvat kuitenkin pitkälti maantieteellisiin alueisiin, ekosysteemeihin, ekologisiin yhteisöihin ja lajeihin, mutta geneettiseen monimuotoisuuteen ja laji-populaatiojatkumoon ei kiinnitetä niin paljon huomiota. Suurin osa luonnonsuojelun harjoittajista keskittyykin lajeihin biologisen monimuotoisuuden mittarina ja tarkastelee jonkin verran intensiivisesti hoidettujen uhanalaisten lajien sisäisiä erityisyksiköitä. Tässä yhteydessä halutaan käyttää yksinkertaisia luokitteluja-nimettyjä lajeja ja muita nimettyjä taksoneita (eli alalajeja) ja joskus suojeluyksiköitä lajien sisällä. Erityisesti lajit tarjoavat yhteisen biologisen monimuotoisuuden mittarin, joka tukee monia tieteellisiä ja hallinnollisia pyrkimyksiä (Mace, 2004; Wilson, 2017). Suojelun perusyksikkönä lajeja käytetään usein biologisen monimuotoisuuden arvon määrittämiseen luetteloiden avulla suojelualueiden lajeista, uhanalaisten lajien tunnistamiseksi lainkäyttöalueilla ja bioturvallisuusmenettelyjen pohjana. Lisäksi lajit ovat tyypillisesti tahoja, joiden kanssa tiedotamme suojeluasioista yleisölle.
tämä sysäys yksilöidä luonteeltaan erillisiä yksiköitä on ristiriidassa useiden toisiinsa liittyvien realiteettien kanssa. Ensinnäkään lajiutuminen ei tapahdu hetkessä. Pikemminkin se on tyypillisesti pitkittynyt prosessi (Dynesius and Jansson, 2014; Roux et al., 2016), etenevät eristyneistä populaatioista, joihin kohdistuu ehkä erilaisia valintapaineita, yksiselitteisiksi lajeiksi pitkän ajan kuluessa, usein ympäristön muutoksen tai yksinkertaisen geneettisen sulautumisen aiheuttamina vaihteluväleinä (Kuva 1; Seehausen et al., 2008; Rosenblum et al., 2012). Toiseksi useimmat tunnistetut lajit eivät ole geneettisesti yhdenmukaisia. Divergenssi (drift and longed isolation) ja retikulaatio (geneflow, hybridization, introgression, and rekombination) muokkaavat tehokkaasti maantieteellistä ja genomista vaihtelua lajien sisällä (Cutter and Payseur, 2013; Edwards et al., 2016)ja jotkut lajit voidaan jäsentää pitkiksi eristetyiksi populaatioiksi, joista osa voisi oikeuttaa tunnustamaan ne täydellisiksi mutta ehkä morfologisesti arvoituksellisiksi lajeiksi (Bickford et al., 2007; Jorger ja Schrödl, 2013). Kolmanneksi, niin kauan tunnustettu, nimetty lajit eivät vastaa, kuinka paljon evolutionary perintö ne edustavat-yksinkertaisesti sanottuna vesinokkaeläin (edustajana muinoin eriytynyt sukua) voidaan pitää arvokkaampi kuin äskettäin eriytynyt jyrsijä (Isaac et al., 2007). Neljänneksi, lajinimitykset eivät ole kiveen heitettyjä, vaan ne ovat hypoteeseja, joita on testattava uusien todisteiden tullessa saataville (Hey et al., 2003). Jokainen näistä kysymyksistä aiheuttaa monimutkaisuutta ja epäselvyyttä lajien yleisessä rajaamisessa ja suojeluhuomion osoittamisessa niille tai niiden osapopulaatioille. Lisäksi lajien rajaamiseen liittyy myös erilaisia lajikäsityksiä ja niiden diagnosointitapoja, mikä on aiheuttanut suurta tyrmistystä suojelubiologien keskuudessa (Mace, 2004; Frankham et al., 2012; Garnett ja Christidis, 2017).
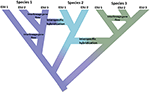
kuva 1. Lajiutuminen jatkumona, joka etenee eristyneistä populaatioista yksiselitteisiksi lajeiksi, mikä näkyy värimuutoksena eri lajeja edustavien värien kanssa. Jotkin populaation eristyneisyydestä johtuvat sukusolut kehittyvät oikeiksi lajeiksi, joiden puun sisäiset solmut edustavat lajiutumisprosessin käynnistymistä. Muut sukuhaarat eivät kehity varsinaisiksi lajeiksi, vaan eroavat toisistaan ajan myötä osoittaen, etteivät lajit ole geneettisesti yhtenäisiä. Nämä lineaarit esitetään Evolutionaarisesti merkittävinä yksikköinä (ESU). Sukujen välinen geenivirta on osoitettu lajeille 1 ja lajeille 3. Tällöin evolutiivinen riippumattomuus voi olla ohimenevää, koska kaksi samaan lajiin kuuluvaa sukua eivät ole lisääntymiskykyisesti eristyksissä. Vaikka ne pysyvät erillisinä sukulinjoina (lajeina), lajienvälinen hybridisaatio voi tapahtua rajoitetulla geeninvaihdolla (värinmuutoksella) lajien välillä.
sellaisten tehokkaiden suojelustrategioiden toteuttaminen, joilla pyritään suojelemaan sekä lajeja että geneettistä monimuotoisuutta, hyötyisi sekä johdonmukaisemmasta näkemyksestä siitä, miten rajaamme lajit ja muut yksiköt populaatio-lajijatkumossa, että siitä, miten näitä yksiköitä tulkitaan eri lainsäädännöissä maailmanlaajuisesti. Genomiikan uudet kyvyt tarjoavat sekä mahdollisuuksia että haasteita näiden tavoitteiden saavuttamiseen. Seuraavassa tarkastelemme: i) miten genomimittakaavaiset analyysit voidaan sovittaa yhteen varovaisen lähestymistavan kanssa lajien rajaamisessa ja nimeämisessä, samalla kun yksilöidään lajien geneettisen monimuotoisuuden tärkeimmät osatekijät, ii) miten voidaan käyttää useiden maanosien voimassa olevaa suojelulainsäädäntöä, jossa ”lajit” määritellään niin, että niihin sisältyy laaja valikoima infrakohtaisia luokkia, kun otetaan huomioon genomiseulojen tarjoama korkea resoluutio, ja iii) miten lajiutumisprosesseja ja genomiikan vaihtelun jäsentämistä koskevat oivallukset ovat yhdenmukaisia lainsäädännön ja sen jälkeen hallinnoinnin kanssa. Ehdotamme sitten joitakin keinoja, joilla suojeluyksiköitä yhdenmukaistetaan lainsäädännöllisten lähestymistapojen kanssa, jotta voidaan suojella koko monimuotoisuutta populaatioilta lajien kautta.
genomiikka, lajiutuminen ja taksonomia
monien biologisten tieteenalojen tavoin systematiikka, taksonien nimeämiskäytäntö sekä niiden suhteiden ja niiden muodon ymmärtäminen edistyvät merkittävästi genomiikan ja tilastollisten mallien avulla. Onko kautta koko genomin sekvensointi (esim., Nater et al., 2017) tai genominäytteenotto (SNPs, target capture-sequencing; Jones and Good, 2016; Leache and Oaks, 2017), meillä on nyt ennennäkemätön resoluutio malleja geneettisen monimuotoisuuden (Funk et al., 2012). Yhdessä yhä tehokkaampien tilastollisten mallien kanssa, joilla voidaan päätellä perimän ja lajien eroavaisuuksia, nämä tiedot tarjoavat uusia oivalluksia evolutionaarisista prosesseista, jotka synnyttävät lajeja ja geneettistä vaihtelua.
näiden uusien työkalujen soveltaminen tuottaa kaksi vastakkaista näkemystä. Toisaalta monet tutkimukset paljastavat hyvin erilaisia geneettisiä populaatioita nimetyissä lajeissa, niin kutsutuissa kryptisissä lajeissa (Bickford et al., 2007; Jorger ja Schrödl, 2013; Struck et al., 2017). Tämä ei ole mitään uutta, koska se perustuu pitkään historiaan spatiaalisen populaatiogenetiikan ja molekyylifylogeografian, joka on ollut tuottava pyrkimys kolme vuosikymmentä (Avise et al., 1987). Mitä genomiikka tuo tähän pyrkimykseen on sekä väestörakenteen lisääntynyt resoluutio että tiedot, jotka riittävät tilastollisesti päättelemään väestöhistorioita ja testaamaan vaihtoehtoisia eroavuusmalleja (Degnan and Rosenberg, 2009; Sukumaran and Knowles, 2017). Toisaalta genomimittakaava-analyysit paljastavat myös, että (niin kauan kuin kasveja on tunnustettu), eläinlajien geenivaihto (introgressio) on yleisempää kuin aiemmin luultiin, sekä lajiutumisen aikana että sen jälkeen, ja se voi jopa ajaa uusia adaptiivisia säteilyjä (Mallet, 2007; Rieseberg and Willis, 2007; Arnold and Kunte, 2017) Kuva 1. Yhdessä nämä oivallukset korostavat edelleen lajiutumisen pitkittymistä (Roux et al., 2016) ja paljastavat fylogeografia-fylogenetiikka-jatkumon kautta toimivat vastavoimat (Edwards et al., 2016).
miten tämä kaikki liittyy taksonomiaan ja suojelutavoitteisiin? On olemassa vahva ja perusteltu huoli siitä, että lisääntynyt resoluutio tarjoama genomiseuloja voisi johtaa rehottava yli-jakaminen lajien (Isaac et al., 2004), mahdollisesti rajoittamalla hallinnoinnin joustavuutta ja saattamalla pienet geneettisesti poikkeavat populaatiot sisäsiittoisuuteen ja lopulta sukupuuttoon (Frankham et al., 2012). Päinvastoin, Gippoliti et al. (2018) väittävät, että taksonominen inertia, mikä aiemmin ei tunnista monia lajeja Afrikan sorkka-ja kavioeläinten, oli johtanut outbreeding masennus sekoittumisen jälkeen ja epäonnistuminen suojella maantieteellisesti rajoitettu taksonomia. Heidän näkemyksensä (ja katso myös Morrison et al., 2009)on, että ei ole näyttöä siitä, että taksonominen inflaatio vaikuttaa kielteisesti sorkka-ja kavioeläinten suojeluun (Gippoliti et al., 2018). Tähän liittyy huoli siitä, että epävakaa taksonomia hidastaa sekä konservointikäytäntöä että yleisiä käsityksiä (Garnett and Christidis, 2017). Nämä huolenaiheet johtuvat osittain systemaattisen biologian lajikäsitteistä ja diagnostisista menetelmistä käydystä pitkään jatkuneesta keskustelusta, erityisesti fylogeneettisen Lajikäsitteen soveltamisesta, jonka mukaan lajit ovat minimaalisesti diagnosoitavissa olevia yksiköitä (Cracraft, 1983). Kuitenkin suurin osa taksonomit ovat luonnostaan konservatiivinen, vain nimeämällä lajeja, kun on concordance poikki useita linjoja todisteita (Dayrat, 2005; Padial et al., 2010; Schlick-Steiner ym., 2010: Yeates et al., 2011). Meidän näkemyksemme on, että lajien tulisi edustaa vankkaa itsenäisesti kehittyvää sukua, joka säilyy suurelta osin koskemattomana ollessaan kosketuksissa lähisukulaisiin—Evoluutiolajikäsitteen (Wiley, 1978) ja yleistetyn Perimälajikäsitteen (de Queiroz, 1998, 2007) tarkoituksen mukaisesti. Käytännössä tämä edellyttää suoraa näyttöä lisääntymis-eristäytymisestä tai kohtuullisesta sijaissynnytyksestä samalla kun sallitaan rajoitettu geenivaihto (Singhal et al., 2018).
Genomitiedot antavat aiempaa suuremman kyvyn tunnistaa eriytyminen ja erilaistuminen lajien sisällä ja niiden välillä, mikä edellyttää suurempaa pohdintaa siitä, miten tämä vaikuttaa taksonomisten yksiköiden rajaamiseen. Jos populaatiot ovat sekä geneettisesti että fenotyypillisesti hyvin erilaisia jo nimettyihin lajeihin verrattuna, on näiden nimeämiselle erillisiksi lajeiksi vahvat perusteet. Jos populaatiot eroavat voimakkaasti fenotyypiltään ja niiden genominen on vähäistä, on arvioitava, ovatko nämä fenotyyppiset erot I) periytyviä ja ii) aiheuttavatko ne merkittävää lisääntymisterveyttä. Genomikuvaukset voivat ratkaista tällaiset tilanteet testaamalla rajoitettua geenivirtaa. Haastavampaa on, missä havaitaan morfologisesti samanlaisia, mutta geneettisesti toisistaan poikkeavia populaatioita. Tällaiset populaatiot voidaan helposti diagnosoida erillisiksi evolutiivisiksi linjoiksi soveltamalla tilastollisia karsintamenetelmiä multilocus-aineistoon (Carstens et al., 2013; Rannala, 2015), mutta ovatko ne ohimeneviä populaatioita vai kestäviä lajeja (Sukumaran and Knowles, 2017)? On varmasti mahdollista, sisällä sääntöjä nimikkeistö, tunnistaa lajien DNA sekvenssit yksin, tarjoamalla sekvenssipohjainen diagnostiikka (Tautz et al., 2003; Cook ym., 2010). Esimerkiksi Murphy et al. (2015) rajaa ja kuvaa kuusi australialaista aavikkosukupolvilajia DNA-sekvenssitietojen perusteella huolimatta selkeistä morfologisista eroista. Tämä on kuitenkin yleinen parannettava alue, jossa systematistien on sovellettava termiä ”kryptiset lajit”johdonmukaisemmin ja täsmällisemmin (Struck et al., 2017).
tilanteissa, joissa DNA-sekvenssitiedot tarjoavat potentiaalisen keskeisen ominaisuuden lajin tunnistamiseksi, ehdotamme, että on erityisen tärkeää soveltaa muita todistusviivoja, jotta vältetään ylijännittyminen (Oliver et al., 2015). Yksi lähestymistapa on testata huomattavaa lisääntymisterveyttä silloin, kun tällaiset populaatiot muodostavat luonnollisia kosketusvyöhykkeitä. Esimerkiksi vertailevassa genomianalyysissä Kontaktivyöhykkeistä Carlia-liskojen sukulinjojen välillä, jotka ovat fenotyyppisesti erottamattomia, geneettisen eristyneisyyden indeksit korreloivat voimakkaasti divergenssiajan kanssa (Singhal and Moritz, 2013; Singhal and Bi, 2017) (Kuva 2). Enemmän eriäviä näiden sukujen-paria on nyt virallisesti tunnustettu erillisiksi lajeiksi, kun taas lähempää sukua (vielä tilastollisesti diagnostisoitavissa) sukulinjoja, joilla on näyttöä genomisesti laaja geneettinen vaihto yli kontaktialueiden eivät olleet (Singhal et al., 2018). Koska näiden taksonien divergenssin ja lisääntymis-eristyneisyyden välillä on vahva empiirinen suhde, nämä kirjoittajat erottivat edelleen kaksi (läheistä sukua olevan lajin) allopatrista ja kryptistä isolaattia, joilla oli vielä suurempi genominen divergenssi kuin edellä kuvatuilla. Laajentamalla tämä lisääntymis eristäminen informoitu metriikka muihin kongeneerisiin lajeihin, joilla on syvästi poikkeava mutta morfologisesti samanlainen fylogeographic lineages (Potter et al., 2016, 2018; Afonso Silva et al., 2017a) viittaa siihen, että lisää lajeja on vielä kuvattavana, erityisesti saarilla (kuva 2, Carlai amax ja C. rufilatus), ja joitakin tällaisia lajeja on kuvattu täydentävän fenotyyppianalyysin perusteella (Afonso Silva et al., 2017b). Muut sukulinjat, joilla on uudempia eroja, mutta jotka on silti tilastollisesti rajattu erillisiksi sukulinjoiksi käyttäen suurta joukkoa ydingeenejä, voidaan esittää Evolutionaarisesti merkittävinä yksikköinä (ESU) lajien sisällä.
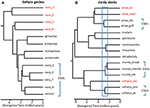
kuva 2. Fylogenomic resolution of species borders in lizards from the Australian Monsoonal Tropics (AMT). Kussakin tapauksessa Relaatiot ja suhteelliset eroajat on arvioitu 100 eksonista käyttäen tähtipeite2: ssa toteutettua monilajikoalitiota, mutta näiden kahden esimerkin väliset aikaskaalat eivät välttämättä ole samat. (A) Gehyra geckojen nana-ryhmän tulokset (Moritz et al., 2018). G. Nanan suvun fylogeneettiseen monimuotoisuuteen, kuten aiemmin todettiin, kuului neljä muuta jo kuvattua ja morfologisesti erottuvaa lajia. Myöhemmät analyysit vaihteluvälien päällekkäisyyksistä ja morfologiasta ovat johtaneet siihen, että kukin fylogeneettisesti eriytyvistä linjoista (punaiset taksonimerkit) on tunnistettu erillisiksi lajeiksi (Doughty et al., 2018). G. Nanassa jäljellä olevia (morfologisesti erottamattomia) sukulinjoja, sellaisina kuin ne nyt tunnustetaan, voidaan pitää ESUs: Ina. (B) lajien rajojen jatkuva selvittäminen Carlia skinksissä AMT: ltä. Jotkin sukusolut on jo tunnustettu lajeiksi geneettisen lajirajauksen ja diagnostisten morfologisten erojen jälkitunnistuksen (C. insularis ja C. isostriacantha; Afonso Silva ym., 2017b). Muille komplekseille (Potter et al., 2016, 2018), taksonomiset korjaukset ovat käynnissä. Sininen suorakulmio osoittaa divergenssiaikojen vaihteluvälin, jossa on havaittu huomattavaa lisääntymis-eristyneisyyttä, joka korreloi divergenssin kanssa, morfologisesti kryptisten sukulaislajien välisten kontaktivyöhykkeiden välillä (Singhal and Bi, 2017). Konservatiivisesti ja seuraten (Singhal et al., 2018), tunnistamme ehdokaslajeiksi kolme geneettistä sukulinjaa, joiden erot ylittävät tämän empiirisen kynnyksen. Tämä jättäisi geneettisesti erilliset ESUs: t jokaiseen Carlia amaxiin, Carlia mundaan ja Carlia rufilatukseen (siniset sulut). Vaa ’ at edustavat arvioita eroavaisuuksien ajasta miljoonissa vuosissa.
ääritapauksissa genomimenetelmät paljastavat yksittäisten taksonomisten lajien olevan paitsi geneettisesti heterogeenisia, myös fylogeneettisessä mielessä myös muita nimettyjä lajeja. Tällainen ”parafyly” voi syntyä, vaikka viimeaikainen lajiutuminen yhdestä, geneettisesti eriytyneestä lajista (Patton and Smith, 1994), mutta se luo myös hypoteesin, että geneettisesti heterogeeninen taksoni voi edustaa useita lajeja. Jälleen tarvitaan lisää todisteita. Esimerkkinä voidaan mainita Pohjois-Australiasta peräisin olevien gehyra-gekkoliskojen kompleksin fylogenominenanalyysi, jossa rajattiin yhdeksän evolutionaarista sukulinjaa, joiden joukossa oli neljä jo fylogeneettisesti nimettyä lajia (Moritz et al., 2018; Kuva 2). Myöhemmät todisteet maantieteellisestä päällekkäisyydestä (säilyttäen kuitenkin fylogeneettisen erottelun) ja hienovaraiset morfologiset erot tukivat neljän fylogeneettisesti poikkeavan sukulinjan nimeämistä (Doughty et al., 2018). Vastaavasti lähempää sukua olevilla (mutta tilastollisesti erotettavissa olevilla) sukuhaaroilla oli päällekkäisten jakaumien sijaan vierekkäisiä jakaumia, mutta ne eivät olleet morfologisesti diagnosoitavissa, joten ne säilyivät yhtenä lajina (G. nana sensu stricto), joskin niillä oli useita ESU: ita, lisätutkimuksia odoteltaessa (kuva 2). Myöhemmin näiden ESU: iden rajojen yli otettujen näytteiden genomikuvaukset paljastivat, että ainakin joissakin on merkittävää geneettistä vaihtoa, mikä vahvisti alkuperäisen, konservatiivisen lähestymistavan, jolla lajit tunnistettiin.
muissa tapauksissa kasvien genomitutkimukset, jopa SNP: n yksinkertaisen ordinaation avulla, voivat paljastaa karuja ja ennakoimattomia eroja populaationäytteiden välillä, mikä viittaa kryptisten taksonien esiintymiseen, joita voidaan sitten analysoida diagnostisten fenotyyppisten merkkien varalta. Esimerkiksi SNP-pohjaiset todisteet populaatioiden genomieroista ja Ekotyypin eroista Eukalyptussalubriksessa paljastivat kaksi erillistä molekyylilinjaa, jotka säilyttivät erottuvuutensa maantieteellisellä päällekkäisyydellä, ja ekotyypin sopeutumista pidettiin tärkeänä tekijänä minimoitaessa geenivirtaa sukujen välillä. (Steane et al., 2015; kuva 3). Toisessa tapauksessa, Australian Pelargonium, genomianalyysi yhdistettynä morfologiset ja lisääntymistutkimukset, on johtanut merkittäviin muutoksiin taksonomia, sekä halkaisu ja niputtaminen taksonomia, erityisesti jos aiemmin käytetyt morfologiset piirteet on todettu labiili ja epäsovun kanssa fylogeny (Nicotra et al., 2016).
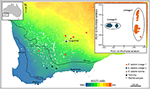
kuva 3. Genomianalyysissä SNP: n tietojen avulla tunnistettiin kryptisiä sukujuuria laajalle levinneessä Eucalyptus salubriksessa. Kunkin sukulinjan näytepopulaatioiden sijainti on esitetty kuivuusindeksigradientin päällä Lounais-Australiassa. 16 122 DArTseq-merkistä johdettuihin binäärisiin geneettisiin etäisyyksiin perustuva PCoA osoittaa kaksi erillistä sukulinjaa. Mukautettu Steane et al. (2015).
edellä mainitut tapaukset kuvaavat integratiivisessa taksonomiassa (Padial et al., 2010; Yeates et al., 2011). Lisäksi nämä tapaukset käyttävät jonkinlaista genomin vähentäminen sekvensointi saada laadukkaita tietoja 1000: n riippumattomien geenien, antaa paljon enemmän luottamusta tuloksiin kuin on kyse käyttämällä vain plastid loci tai muutamia ydingeenejä. Esimerkki siitä, miten integratiivisen taksonomian soveltaminen voi dramaattisesti muuttaa käsityksiä monimuotoisuudesta, on yhdistetty Madagaskarilta peräisin olevien sammakoiden geneettinen, Akustinen ja morfologinen tutkimus, joka lähes kaksinkertaisti tunnetun lajiston monimuotoisuuden (Vieites et al., 2009). Nämä kirjoittajat ehdottivat useita termejä, jotka heijastavat taksonomisen resoluution ja kongruenssin tilaa: Vahvistetut geneettisesti ja fenotyyppisesti poikkeavat lajit, joita ei ole vielä nimetty; vahvistamattomat lajit geneettisesti eriytyville taksoneille, joiden morfologiaa ja akustiikkaa ei ollut vielä tutkittu; ja erittäin selvät sukulinjat yhteisöille, joiden tiedetään olevan geneettisesti erilaisia mutta fenotyyppisesti arvoituksellisia. Genomianalyysejä ei ole vielä sovellettu näihin taksoneihin, mutta se informoisi varmasti päätöksiä salaliittolajeista vs. ehdokaslajeista.
genomiset lähestymistavat johtavat epäilemättä morfologisesti konservatiivisten taksonien jakautumiseen edelleen, vaikka niitä sovellettaisiinkin konservatiivisesti. Mutta päinvastoin, on olemassa selkeitä esimerkkejä, joissa samat lähestymistavat paljastavat yli-split tai väärin diagnosoitu lajeja (Nicotra et al., 2016). Toki kasvi-ja eläinlajikomplekseista on jo olemassa monia molekyylitutkimuksia, jotka perustuvat pienemmän mittakaavan aineistoihin (esim., mikrosatelliitti, mtDNA ja cpDNA), jotka ovat tunnistaneet, missä lajit on diagnosoitu väärin tai missä useita lajeja on rajattu taksonomisesti, mutta niitä pidetään nykyään yhtenä lajina. Esimerkiksi Rabosky et al. (2014) käytti mitokondrio-ja ydingeenifylogenioita scincid ctenotus inornatus-kompleksin tarkistamiseen, mikä johti lajien nettovähennykseen ja lajien rajojen uudelleensuuntaamiseen; aiemmin käytetyt morfologiset piirteet, vaikka ne sopivat kenttäbiologeille, olivat erittäin labiileja ja suurelta osin eivät vastanneet lajien rajoja diagnosoidulla geneettisesti. Kasveissa joitakin tunnetuimpia esimerkkejä löytyy orkideat, kuten Euroopan orkidea suku Ophrys ja Australian orkidea suku Caladenia, jossa molekyylitutkimukset osoittavat huomattavasti vähemmän yhteisöjä, jotka voitaisiin tunnistaa lajiksi, kuin ne perustuvat morfologiset piirteet ja pseudokoopulatorinen pölytysoireyhtymiä (KS.Devey et al., 2008; Swarts et al., 2014).
Genomiasteikkoaineisto, kun sitä tulkitaan konservatiivisesti ja ihanteellisesti fenotyyppien rinnakkaisanalyysien avulla, johtaa edelleen taksonomisiin muutoksiin erityisesti morfologisesti konservatiivisissa taksoneissa (Fišer et al., 2018). Mutta toisin kuin joidenkin esittämät huolenaiheet (Garnett ja Christidis, 2017), katsomme, että tämä on oikea pyrkimys evolutiiviseen systematiikkaan, vaikka tulokset ovat hetkellisesti hankalia.
genomiikka ja Intraspecific Conservation Units
vaikkakin vähemmän kiistanalainen kuin taksonominen rajaus ja lajitason keskustelu, mutta monin tavoin yhtä monimutkainen, on ollut merkittävä keskustelu suojeluyksiköiden määrittelystä ja rajaamisesta tärkeinä intraspecific diversity-tekijöinä, jotka on otettava huomioon suojelutoimissa (Ryder, 1986; Waples, 1991; Dizon et al., 1992; Moritz, 1994; Vogler ja de Salle, 1994; Crandall et al., 2000; Fraser and Bernatchez, 2001; Avise, 2004; Funk et al., 2012) (kuva 4). Ottaen huomioon varovainen lähestymistapa tunnistaa lajien genomisesta näyttöä, että me puoltaa edellä (ja katso myös Oliver et al., 2015; Singhal et al., 2018), on väistämätöntä, että joidenkin taksonomisten lajien sisällä on vielä pitkiä eristäytyneitä populaatioita. Sellaisia käsitteitä kuin Evolutionaarisesti merkittävät yksiköt (ESUs) tai Mitoitettavat yksiköt (dus) on ehdotettu täydentämään olemassa olevaa taksonomiaa (Ryder, 1986; Waples, 1991; Moritz, 1994; Mee et al., 2015). Kuten lajeillakin, tulkinnat ja kriteerit vaihtelevat, mutta yhteisenä tarkoituksena on tunnistaa suojelutoimien kannalta erityisen monimuotoisuuden tärkeimmät osatekijät (Funk ym., 2012). Moritz (1994, 2002) kannatti yksinkertaistavaa mutta käytännöllistä molekylaarista lähestymistapaa yksilöidäkseen itsenäisesti kehittyvät lajinsisäiset segmentit, monimuotoisuuden ”historiallisen” akselin; toiset taas painottavat enemmän ”adaptiivista” akselia, joka sisältää toiminnallisen monimuotoisuuden (Ryder, 1986; Crandall et al., 2000; Fraser and Bernatchez, 2001; Waples et al., 2001; Mee et al., 2015) (kuva 4). Riippumatta näiden vaihtoehtojen suhteellisista ansioista on selvää, että populaatiogenomiikan teknisillä ja analyyttisillä välineillä voidaan nyt selvästi tunnistaa molemmat intraspesifisen monimuotoisuuden osatekijät (Funk ym., 2012; Hoban et al., 2013; Whitlock, 2014; Yang ja Rannala, 2014).
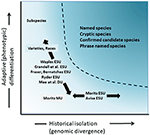
kuva 4. Geneettisen monimuotoisuuden erottaminen kahteen osaan: Adaptiivinen vaihtelu, joka syntyy luonnonvalinnan kautta, ja analysoidaan vaikka fenotyyppien analyysi ja historiallinen eristyneisyys tai neutraali divergenssi, joka arvioidaan genomisen divergenssin kautta. Käyrän yläpuolella oleva tummempi varjostettu alue osoittaa olosuhteet, joissa populaatioita todennäköisesti pidetään erillisinä lajeina useimmissa käsitteissä. Erilaisia suojeluyksikkökäsitteitä ja infrakohtaisia taksonomisia kategorioita esitetään suhteessa geneettisen monimuotoisuuden kahteen akseliin (Moritzin jälkeen, 2002).
Tämä johtaa kysymykseen: miksi ei vain nimettäisi esuksia alalajeiksi? Sekä kasvi-että eläintieteilijät ovat perinteisesti määritelleet ja pitäneet alalajeja fenotyyppisesti erillisinä, allopatrisina populaatioina, jotka voivat risteytyä toisiinsa maantieteellisillä rajoilla (Mayr, 1942; Grant, 1981). Sellaisenaan ne mahdollisesti heijastavat adaptiivista eroavuutta ilman, että ne ovat välttämättä olleet historiallisesti eristyksissä. Alalajinimityksiä käytetään laajalti kasveissa, ja maantieteelliset ja ekologiset erot ovat ensisijainen tapa erottaa ne (Hamilton and Reichard, 1992). Siltä osin kuin alalajit ilmentävät paikallista sopeutumista joko jatkuvalla geenivirralla tai ilman sitä, ehdotamme, että fenotyyppisesti määritellyt alalajit olisi edelleen tunnustettava suojelussa eikä yhdistettävä ESU: iden kanssa, ainakaan ESU: iden, jotka on määritelty edustamaan historiallisesti eristyneitä populaatioita.
samalla tavalla kuin populaatiogenomiikka tarjoaa paljon suuremman tarkkuuden ESU: iden tunnistamisessa käyttämällä sekä neutraaleja että adaptiivisia lokuksia, se tarjoaa myös keinon käyttää suuria määriä neutraaleja lokuksia helpommin rajattaviin Hallintayksiköihin (MUs) demografisesti riippumattomina yksikköinä, joille on ominaista rajoitettu geenivirta (Funk et al., 2012). Vaikka suurin osa keskitymme tässä ESU: n ja vastaaviin yksiköihin, kuten du: hun, jotka voidaan tunnustaa tärkeimmiksi erityisten suojeluyksiköiksi ja jotka on hyväksytty suojeltaviksi joillakin lainkäyttöalueilla, MUs: ien on myös tunnistettu edustavan demografisesti itsenäisiä populaatiokokonaisuuksia(KS. Palsbøll et al., 2007), joita voidaan hallita säilyttämään suurempi ESU (Moritz, 1994; Funk et al., 2012). Hallintoyksiköt voivat olla tärkeitä lajien populaatioiden seurannassa ja hallinnassa, ja niitä on kutsuttu myös ”kalakannoiksi”, jotka ovat erityisen merkityksellisiä kalastuksenhoidon kannalta (Avise, 2004; Laikre et al., 2005). Moritzin (1994) mus-määritelmä ”populaatioiksi, joilla on merkittäviä eroja alleelien taajuuksissa ydin-tai mitokondrio-lokuksissa riippumatta alleelien fylogeneettisestä erottuvuudesta” on yleisesti hyväksytty, vaikka, kuten palsbøll ym. (2007), demografinen yhteys on avaintekijä, ei historiallisen geenivirran taso.
suojeluyksiköiden rajaamisella erityisen monimuotoisuuden tärkeinä osatekijöinä on myös merkittäviä vaikutuksia pienten sisäsiittoisten populaatioiden geneettiseen pelastamiseen geenivirran ennallistamisen tai lisäämisen kautta, ja on yhä selvempää, että geneettistä pelastamista on tarkasteltava laajemmin, jotta lisääntyneet populaatiosukupuutot voidaan estää (Love Stowell et al., 2017; Ralls et al., 2018). Moritz (1999) ehdotti, että MUs: ien mutta ei ESUs: ien sekoittamista voitaisiin pitää sopivana strategiana geneettiselle pelastukselle. Kuten korostivat Frankham et al. (2012) hallinnoijat eivät yleensä pitäisi taksonomisesti tunnustettujen lajien sekoittamista, jolloin taksonominen liikakalastus voisi rajoittaa mahdollisuuksia pelastaa pieniä sisäsiittoisia populaatioita. Ehdotamme, että vaikka tiukempi lähestymistapa tiettyjen yksiköiden, kuten MUs: n, rajaamiseen voisi helpottaa tätä ongelmaa, on myös selvää, että siirtymisnopeuksien ja adaptiivisen divergenssin genomiarvioinnilla on mahdollisuus tarjota paljon vankempia ohjeita paitsi lajirajojen ja intraspesifisen vaihtelun määrittelyssä myös geneettisten pelastusstrategioiden suunnittelussa.
Fylogenomiikka ja Konservointi
genomiikka yhdistettynä fylogeneettisiin menetelmiin voi helposti ratkaista taksonien välisiä suhteita ja evolutiivista etäisyyttä sekä rajata lajeja. Tämä saavutetaan parhaiten käyttämällä lajipuu menetelmiä, eikä concatenation, koska entinen huomioon geenipuun heterogeenisuus ja välttää yli-inflaatio kärjen pituudet johtuu esivanhempien polymorfismi (Edwards et al., 2016; Ogilvie et al., 2016). Vaikka massiivinen määrä loci eivät useinkaan ole välttämättömiä, myös enemmän loci usein parantaa resoluutio sekä suhteita ja haara pituudet (esim.Blom et al., 2016). Suojelun merkitys on kaksijakoinen. Ensinnäkin fylogeneettisten arviointien avulla voidaan tarkastella lajien tai ESUs: ien erottamiskykyä, ei vain sitä, kuvataanko niitä. Niinpä EDGE-ohjelma (Isaac et al., 2007) kannattaa lajien painottamista suojeluun sen perusteella, kuinka ainutlaatuista evoluutiohistoriaa ne edustavat tavanomaisten IUCN: n kriteerien lisäksi. Toiseksi, oksien pituudet itsessään voivat edustaa biodiversiteettiarvoja riippumatta taksonomiasta (Faith, 1992), joka on osoittanut arvoa suojelualueiden priorisoinnissa (Maally et al., 2015; Rosauer et al., 2016, 2018). Tässä mielessä fylogeneettisten mittareiden omaksuminen fylogeneettisen monimuotoisuuden ja fylogeneettisen endeemisyyden kaltaisilla toimenpiteillä mahdollistaa sen, että voidaan korostaa erittäin evolutiivisen monimuotoisuuden alueita, jotka voidaan kohdentaa suojeluun ja jotka asetetaan etusijalle kaikissa biologisen monimuotoisuuden suojelun parantamista koskevissa suunnitteluprosesseissa. Tämä lähestymistapa voi parantaa geneettisen monimuotoisuuden ja evolutionaaristen prosessien suojelua eri eliöryhmille eri maantieteellisillä alueilla (Maally et al., 2015) päätösten tekeminen alueista, joilla priorisoidaan vahvemmin oikut siitä, miten genomiset todisteet käännetään lajien tai tiettyjen yksikköjen tunnistamiseksi.
biologisen monimuotoisuuden säilyttämistä koskevat reaalimaailman lainsäädännön määritelmät
politiikka ja lainsäädäntökehykset vaihtelevat kansainvälisistä yleissopimuksista ja strategioista kansallisten strategioiden ja lainsäädännön kautta sekä valtioiden tai alueiden lainsäädännöstä ja strategioista maiden sisällä. Tärkeimpiä biologista suojelua koskevia kansainvälisiä välineitä ovat osapuolten yleissopimuksella (COP) täytäntöön pantu biologista monimuotoisuutta koskeva yleissopimus, Maailmanperintösopimus, Maailmanlaajuinen Biodiversiteettistrategia, metsien suojelua koskeva Montrealin prosessi, kosteikkoja koskeva Ramsarin yleissopimus, muuttavia lajeja koskeva yleissopimus, uhanalaisten lajien kansainvälistä kauppaa koskeva yleissopimus (CITES), Kansainvälisen luonnonsuojeluliiton (IUCN) uhanalaisia lajeja koskeva punainen luettelo ja ilmastonmuutosta koskeva Yhdistyneiden Kansakuntien puitesopimus. Kaikissa näissä peruskirjoissa lajien ja geneettisen monimuotoisuuden säilyttämisen päämäärä on biologisen monimuotoisuuden suojelun perustavoite, ja lajit ovat laajasti määritelty suojeluyksikkö. Lainsäädännön ja kansainvälisten direktiivien mukaiset lajien määritelmät ovat kuitenkin hyvin vaihtelevia. Niihin sisältyy lajien taksonominen tunnustaminen ja lajin sisäiset taksonit, mutta niihin voi sisältyä myös geneettisiä/evolutiivisia käsitteitä, jotka usein kuuluvat ”lajin” oikeudellisen määritelmän piiriin (Taulukko 1).

Taulukko 1. Maailman Lainsäädäntö ja direktiivit, jotka määrittelevät ”lajit” luetteloitaviksi ja suojeltaviksi.
kun otetaan huomioon suojeltavien infrakohtaisten tasojen laaja kirjo (Taulukko 1), maiden lainsäädännön ja siihen liittyvien luettelointiprosessien pitäisi helpottaa geneettisen monimuotoisuuden säilyttämistä. Esimerkiksi ”lajin” määritelmä, kun tarkastellaan lainsäädäntöä ympäri maailmaa (Taulukko 1), voi sisältää: lajit, alalajit, lajikkeet ja maantieteellisesti ja/tai geneettisesti erilliset populaatiot, kun taas Etelä-Afrikan lainsäädännön mukaan siihen voi kuulua myös lajikkeita, maantieteellisiä rotuja, kantoja ja risteymiä. Näin ollen useimpien lakien mukaan ”laji” sisältää kokonaisuuksia, jotka heijastavat erilaisia infra-spesifisiä tasoja, jotka kaikki ovat osa laajempaa lajijakaumaa ja heijastavat vaihtelevassa määrin populaatio-laji-jatkumoa. Tämä vastaa sitä väistämätöntä vaikeutta, jonka taksonomit, evoluutiobiologit ja luonnonsuojelun geneetikot kohtaavat löytäessään johdonmukaisemman ja yhdenmukaisemman tavan määritellä yksiköt tässä jatkumossa.
jotkut väittävät, että huoli lajien poliittisista vaikutuksista ja infrakohtaisesta epävarmuudesta on aiheeton (Hey et al., 2003), mutta olemme edelleen sitä mieltä, että vaikka lainsäädännöllä voidaan suojella geneettistä monimuotoisuutta eri tasoilla, biologien tehtävänä on selkeyttää ja yhdenmukaistaa sitä, miten nämä tasot olisi rajattava lajeista yksittäisiin populaatioihin. Tällä hetkellä johtajille ja lainsäätäjille jää hämmentävän paljon lajikäsitteitä ja tieteentekijöiden tarjoamia infrayksilöitä, kun populaatio-lajijatkumon yksiköitä rajataan. Esimerkiksi kaikki lainkäyttöalueet pitävät taksonomien tunnistamia lajeja suojelun avainyksikköinä (Taulukko 1), mutta taksonomien keskuudessa käydään edelleen laajaa keskustelua lajikäsitteistä, ja lajien toteaminen käytännössä vaihtelee ryhmien sisällä ja niiden välillä. Tämä on johtanut viimeaikaiseen pyyntöön säännellymmästä lähestymistavasta taksonomiaan (Garnett and Christidis, 2017), johon muut ovat vastanneet, että taksonomian sääntely ei ole vastaus (Thomson et al., 2018). Vaikka olemme samaa mieltä joitakin kohtia Thomson et al. (2018) ehdotamme, että on melko naiivia ehdottaa, että lainsäädäntöä itsessään on muutettava siten, että se viittaa nimenomaisesti nimen tarkoittamaan tiettyyn taksonikäsitteeseen. Näissä ja aiemmissa keskusteluissa korostetaan, että evoluutiobiologien ja taksonomien on näytettävä esimerkkiä selkeyden (avoimuuden) varmistamisessa siinä, miten he muuntavat genomiset ja muut todisteet lajien tai erityisten yksiköiden rajaamiseksi ja miten nämä taksonit liittyvät asiaankuuluvaan säilyttämistä ja suojelua koskevaan lainsäädäntöön.
hoito
kuten edellä on todettu, kaikissa lainsäädännöissä ja direktiiveissä tunnustetaan lajitaso suojelun perusyksiköksi, mutta säädetään myös erilaisten infrakohtaisten yksiköiden tunnustamisesta. Monilla on myös implisiittinen vaatimus lajien viralliselle nimeämiselle ja taksonomiselle kuvaukselle, kun niitä harkitaan suojelun priorisointiin ja muodolliseen suojeluun. Esimerkiksi IUCN: n punaisen luettelon ohjeissa ei sallita nimeämättömien lajien luettelointia ja vaaditaan, että uusi lajikuvaus julkaistaan neljän vuoden kuluessa siitä, kun laji on sisällytetty IUCN: n punaiseen luetteloon tai se poistetaan. Tämä korostaa taksonomisten nimien ja kuvausten tärkeyttä sekä lajeilla että tietyntasoisilla tasoilla, rajallista taksonomista tietoa monille kasvi-ja eläinryhmille (Mace, 2004; Oliver and Lee, 2010; Wilson, 2017) ja tarvetta nopeuttaa taksonomisia ponnistuksia erityisesti biodiversiteettirikkailla alueilla (Riedel et al., 2013; Wege et al., 2015; Wilson, 2017).
taksonomisesti nimetyt lajit ja sisäiset taksonomiset luokat, kuten alalajit ja lajikkeet, antavat yleensä perusyksiköt viralliseen luettelointiin, säilyttämisen priorisointiin ja suojeluun, mutta joillakin lainkäyttöalueilla muut intraspesifisten suojeluyksiköiden luokat tunnustetaan virallisesti. Nämä tunnistetaan yleensä yhdistettyjen genomi-ja fenotyyppitietojen avulla. Esimerkiksi USA: n uhanalaisten lajien laki tunnustaa erilliset Populaatiosegmentit (DPSS), jotka perustuvat laajasti ESUs: iin (Waples, 1991), kun taas Kanadan Species at Risk Act tunnustaa DUs: n. Canadian Species at Risk Act-lain mukaisen ”erillisten populaatioiden” operatiivisen tulkinnan tarkoituksena on tunnistaa erilliset populaatiokokonaisuudet, jotka edustavat lajien evolutionaarisen perinnön tärkeitä osia ja joita ei todennäköisesti korvata luonnollisella hajaannuksella (Mee et al., 2015).
sekä DUs että DPSs ovat käsitteellisesti päällekkäisiä ESUs: ien kanssa (Waples et al., 2013) ja sitä on sovellettu laajimmin Pohjois-Amerikan kaloihin, mukaan lukien Luoteis-Amerikan Lohikalat (WAPLES, 1995) ja Kanadan coregonidit (Mee et al., 2015) mielenkiintoista DPSs Yhdysvalloissa sovelletaan vain selkärankaisia, kun taas Kanadassa DUs voidaan soveltaa mihin tahansa organismiin, joka voidaan luetella sen laki (KS.Waples et al., 2013), jossa DUs on tällä hetkellä luettelossa niveljalkaisten, nilviäisten ja kasvien osalta. Australiassa, kuten Kanadassa, erilliset populaatiot voidaan luetella minkä tahansa organismin osalta, joka on tunnustettu ympäristönsuojelua ja biodiversiteettiä koskevan lain (Environment Protection and Biodiversity Conservation Act 1999, EPBC Act) nojalla, vaikka ne eivät ole erikseen nimetty luokka, kuten DUSs ja DPSs. Toistaiseksi niitä on kuitenkin tunnistettu vain selkärankaisille. Yhdysvalloissa on 127 luonnonsuojeluun tarkoitettua selkärankaisten DPSs: ää (https://ecos.fws.gov/ecp/); Kanadassa on lueteltu 70 du: ta, jotka kattavat kaikki eliöt (http://www.registrelep-sararegistry.gc.ca/species/schedules_e.cfm?id=1); ja Australiassa 7 selkärankaislajin populaatioilla, jotka kattavat nisäkkäitä, kaloja ja lintuja, on tunnustettu olevan erityinen suojelutaso EPBC Act-lain nojalla (http://www.environment.gov.au/cgi-bin/sprat/public/publicthreatenedlist.pl). Vaikka arviot näistä populaatioista Yhdysvalloissa ja Kanadassa käyttävät yleensä geneettistä tietoa (Waples et al., 2013), Australiassa ne perustuvat yleensä maantieteelliseen ja juridiseen tunnistamiseen, vaikka joissakin näistä tapauksista niitä on tuettu nisäkkäiden geneettisen erilaistumisen analyysillä (Firestone et al., 1999; Armstrong, 2006)ja fish (Colgan and Paxton, 1997; Ahonen et al., 2009).
luettelo kasvien ja selkärangattomien erityisistä suojeluyksiköistä Kanadassa (DUs) mutta ei Yhdysvalloissa tai Australiassa korostaa toista seikkaa, joka ei liity erityisesti suojeluyksiköiden rajaamiseen vaan laajemmin merkittäviin epäjohdonmukaisuuksiin biologisen monimuotoisuuden säilyttämisen priorisoinnissa; ikoniset selkärankaiset, kuten linnut, nisäkkäät ja kalat, ovat painopiste monilla lainkäyttöalueilla, kun taas muut organismit eivät. Tämä tiettyjen eliöiden populaatioiden etuoikeutettu luettelointi kuvastaa monia kysymyksiä, jotka eivät liity suojeluyksiköiden rajaamiseen liittyvään tieteeseen vaan pikemminkin populaatioiden luettelointiin yleisen tietoisuuden, poliittisen tarkoituksenmukaisuuden ja luetteloitavan organismin karismaattisuuden perusteella (KS.Waples et al., 2013).
vaikka Australiassa jotkin tunnettujen nisäkkäiden, lintujen ja kalojen populaatiot ovat rauhoitettuja EPBC-lain nojalla, nyt on olemassa huomattava määrä tietoa, joka perustuu matelijoilla, selkärangattomilla ja kasveilla tehtyihin molekyylitutkimuksiin, joissa populaatiot voidaan tunnistaa ESUs: iksi tai MUs: iksi. Huomiota herättäviä esimerkkejä hyvin jäsennellyistä populaatioista ja lajien sisäisistä hyvin erilaisista linjoista, jotka sopisivat helposti ESU: n vaatimukseen, on esimerkiksi tropiikista (kuva 2) ja muualta peräisin olevilla heikosti leviävillä selkärankaisilla. Samoin Australian selkärangattomilla, kuten amfipodeilla ja tuhatjalkaisilla, merkittävä geneettinen rakenne ja fylogeografiset kuviot tukevat diskreettien populaatioryppäiden rajaamista Esus: iksi, jotka ovat maantieteellisesti erilaisia, joilla on kapeat maantieteelliset alueet, jotka rajoittuvat tiettyihin elinympäristöihin ja joita edustaa vain muutama populaatio (Finston et al., 2007; Murphy ym., 2013; Nistelberger et al., 2014a). Usein nämä lyhyen kantaman endeemiset taksonit ovat suojeluun liittyviä huolenaiheita (Harvey et al., 2011) ja sijaitsevat yhdessä endeemism hotspots (esim., Oliver et al., 2017), jotka ovat itse korkeat prioriteetit monen taksonin säilyttämiseen huomiota (Rosauer et al., 2018), Kuten Kimberleyn alue ja Stirlingin alue Lounais-Australiassa.
Australian kasveilla on myös huomattava määrä populaation geneettisiä ja fylogeografisia tutkimuksia, joissa yksilöidään eri sukujen eri lajien eri populaatioryhmiä, jotka heijastavat hajanaisia ja historiallisesti eristyneitä populaatiojärjestelmiä, geologisia ja edafisia komplekseja, ja joilla on vastakkaisia elinympäristöjä kasvillisuuden ja ilmaston kannalta (KS.Broadhurst et al., 2017). Geneettisesti erillisiä populaatioita esiintyy erityisesti lounais-Australian lajeissa (Coates, 2000; Byrne et al., 2014), jossa on useita tutkimuksia, joissa korostetaan merkittävää geneettistä rakennetta ja hyvin erilaisia lineages in a range of plant generays and families including: Acacia (Mimosaceae; Coates, 1988), Atriplex (Amaranthaceae; Shepherd et al., 2015), Banksia (Proteaceae; Coates et al., 2015), Lambertia (Proteaceae; Byrne et al., 2002), Hakea (Proteaceae; Sampson et al., 2014), Eremophila (Scrophulariaceae; Llorens et al., 2015), Pultanaea (Fabaceae; Millar and Byrne, 2013)ja Calothamnus (Myrtaceae; Nistelberger et al., 2014b). Monissa tapauksissa nämä populaatiot tunnustetaan suojelusuunnittelussa valtion virastojen toimesta ja otetaan huomioon ympäristövaikutusten arvioinneissa (KS. Shepherd et al., 2015), vaikka niitä ei ole lueteltu lainsäädännössä suojeltaviksi. Myönnämme kuitenkin, että näiden yksiköiden virallinen tunnustaminen parantaisi niiden säilymistä pitkällä aikavälillä.
nämä australialaiset esimerkit korostavat lisääntyvää tarvetta tunnistaa tämä lajitason alapuolella oleva geneettisen monimuotoisuuden kerros. Haasteena on varmistaa, että hyvin jäsenneltyjä populaatioita ja hyvin erilaisia, suojeluun liittyviä lajeja suojellaan kilpailevien suojeluhuomiota koskevien vaatimusten edessä. Ikonisten lajien lisäksi tämä haaste näkyy myös maailmanlaajuisesti Kanadaa lukuun ottamatta. Jos selkeästi rajattuja suojeluyksiköitä ei suojella, se todennäköisesti lisää kryptisen sukupuuton riskiä ja merkittävän geneettisen monimuotoisuuden häviämistä, mikä voi monissa tapauksissa koskea suvullisia ja historiallisesti eristyneitä populaatioita, jotka ovat säilyneet lajissa miljoonia vuosia (Moritz, 2002).
päätelmät ja suositukset
lajien ja erilaisten infrakohtaisten luokkien luettelointi uhanalaisuuslainsäädännössä tai kansainvälisissä järjestöissä, kuten IUCN: ssä, osoittaa sitoutumista geneettisen monimuotoisuuden suojelemiseen lajien populaatiojatkumossa. Kuitenkin nykyiset lähestymistavat biologisen monimuotoisuuden säilyttämiseen keskittyvät paljolti tämän jatkumon lajipäähän, mutta geneettiseen monimuotoisuuteen ja infrakohtaiseen vaihteluun ei kiinnitetä niin paljon huomiota. Tämä rajoittuneempi keskittyminen lajinsisäiseen geneettiseen vaihteluun viittaa siihen, että vaikka suojelupäälliköt tunnistavat yhä enemmän sen arvon, että geneettistä tietoa otetaan nimenomaisesti huomioon hoitotoimenpiteissä ja priorisoinnissa, heiltä puuttuu usein biodiversiteettitutkijoilta saatavaa toimivaa tietoa (Waples et al., 2008; Laikre et al., 2010; Cook and Sgrò, 2017; Taylor et al., 2017). Tämä tieteen ja säilyttämiskäytännön välinen kuilu vaikeuttaa tehokkaampaa kokonaisvaltaista lähestymistapaa geneettisen monimuotoisuuden säilyttämiseen ja hallintaan.
esitämme, että biodiversiteettitutkijoiden epäselvät näkemykset suojeluyksiköistä perustuvat suurelta osin epäjohdonmukaisuuksiin kahdella tasolla. Ensimmäinen koskee useita lajeja koskevia käsitteitä ja erilaisia diagnostisia menetelmiä, joita käytetään lajien rajaamisessa, mikä aiheuttaa epäjohdonmukaisuutta eri taksonomisten ryhmien välillä. Toinen liittyy taksonomien, evoluutiobiologien ja suojelubiologien epäjohdonmukaisuuteen ja yksimielisyyteen siitä, miten infra-spesifisiä taksoneja ja muita yhteisöjä, kuten ESUs, DUs ja MUs, tulisi kohdella.
vaikka katsomme, että lainsäädäntö on usein jo riittävän joustavaa sen suhteen, miten taksonit (tai ”lajit”) määritellään, katsomme, että populaatiojatkumoon on puututtava järjestelmällisesti ja että suojeluyksiköt on määriteltävä selkeämmin ottaen huomioon sellaiset käsitteet kuin ESU: t ja se, miten ne liittyvät asiaan liittyvään lainsäädäntöön. Korostamme, että tästä kysymyksestä tulee epäilemättä kriittisempi, kun otetaan huomioon genomiikan kyky havaita erilaistuminen asteittain tarkemmassa mittakaavassa ja sen kasvava käyttö lajien ja infra-spesifisten kokonaisuuksien rajaamisessa.
edellisen analyysin perusteella annamme seuraavat suositukset:
(i) genomitiedot tulisi tulkita konservatiivisesti lajeja rajattaessa käyttäen integratiivista taksonomista lähestymistapaa, joka tyypillisesti sisältää pelkkää geneettistä dataa laajempaa näyttöä.
ii) jos geneetikot tunnistavat lajissa useita sukuja tai erillisiä populaatioita, jotka eivät oikeuta lajien tason nostamiseen eivätkä ole taksonomisesti tunnustettuja, on kannustettava voimakkaammin ja yhdenmukaisemmin yksilöimään selkeästi perusteltuja sisäisiä yksiköitä suojelun tason käytännön arviointia tai vaikutusten arviointia varten, ja nämä on nimenomaisesti tunnustettava suojeluyksiköiksi kyseisen lainkäyttöalueen lainsäädännön yhteydessä.
(iii) IUCN harkitsee Conservation Genetics Specialist Group-ryhmänsä ja alueellisten alaryhmiensä kautta parhaiden tieteellisten käytäntöjen mukaisten ohjeiden kehittämistä erityisyksiköiden tunnistamiseksi ja merkitsemiseksi.
(iv) lajien evolutiivisesta genomiikasta ja jatkumosta populaatioista lajeihin syntyvä vivahteikkaampi näkemys pitäisi tutkijoiden viestiä tehokkaammin suojelujohtajille ja poliittisille päättäjille.
laajemmin toistamme viime aikoina useissa arvioissa annettuja suosituksia (Frankham, 2010; Santamaría and Méndez, 2012; Hoban et al., 2013; Moritz ja Potter, 2013; maallikot et al., 2015; Pierson ym., 2016; Cook and Sgrò, 2017; Taylor et al., 2017), jotka korostavat tiedemiesten tarvetta parantaa viestintää ja lisätä tietoisuutta päätöksentekijöiden, valtion virastojen, kansalaisjärjestöjen ja harjoittajien kanssa siitä, että molekyylistystematiikan, molekyylitaksonomian, fylogeografian ja populaatiogeografian tulokset voivat tarjota kriittistä tietoa geneettisen monimuotoisuuden säilyttämisestä ja evoluutioprosessien hallinnasta.
Tekijäosuudet
DC, MB ja CM osallistuivat kaikki käsikirjoituksen laatimiseen ja tarkistamiseen.
Eturistiriitalausunto
kirjoittajat toteavat, että tutkimus tehtiin ilman kaupallisia tai taloudellisia suhteita, joita voitaisiin pitää mahdollisena eturistiriitana.
kiitokset
tätä hanketta tuki Australian Research Council (ARC Linkage Grant No. LP120200063), Australian National University ja Länsi-Australian Department of Biodiversity, Conservation, and Attractions.
Afonso Silva, A. C., Bragg, J. G., Potter, S., Fernandes, C., Coelho, M. M., and Moritz, C. (2017a). Tropical specialist versus climate generalist: diversification and demographic history of sister species of Carlia skinkks from northwestern Australia. Mol. Ecol. 26:4045–4058. doi: 10.1111 / mec.14185
CrossRef Full Text/Google Scholar
Afonso Silva, A. C., Santos, N., Ogilvie, H. A., and Moritz, C. (2017b). Validointi ja kuvaus kahdesta uudesta Luoteis-Australian Rainbow skinkistä, joilla on monilajiset yhteenliittymismenetelmät ja morfologia. PeerJ 5: e3724. doi: 10.7717 / peerj.3724
PubMed Abstract / CrossRef Full Text / Google Scholar
Ahonen, H., Harcourt, R. G., and Stow, A. J. (2009). Ydin-ja mitokondrio-DNA paljastaa uhanalaisten harmaahain populaatioiden (Carcharias taurus) eristyneisyyden. Mol. Ecol. 18, 4409–4421. doi: 10.1111 / j.1365-294x.2009.04377.x
PubMed Abstract | CrossRef Full Text/Google Scholar
Armstrong, K. N. (2006). Phylogeographic structure in Rhinonicteris aurantia (Chiroptera: Hipposideridae): implications for conservation. Acta Chiropterologica 8, 63-81. doi: 10.3161/1733-5329(2006)82.0.CO
CrossRef Full Text / Google Scholar
Arnold, M. L., ja Kunte, K. (2017). Adaptiivinen geneettinen vaihto: sekoittumisen ja evolutionaarisen innovaation sotkuinen historia. Suuntaukset Ecol. Evol. 32, 601–611. doi: 10.1016/J. puu.2017.05.007
PubMed Abstract | CrossRef Full Text/Google Scholar
Avise, J. C. (2004). Molekyylimerkit, luonnonhistoria ja evoluutio. 2nd Edn. Sunderland, MA; Oxford: Oxford University Press.
Google Scholar
Avise, J. C., Arnold, J., Ball, R. M., Bermingham, E., Lamb, T., Neigel, J. E., et al. (1987). Tietynlainen fylogeografia: mitokondriaalinen DNA-silta populaatiogenetiikan ja systematiikan välillä. Ann. Rev. Ecol. Sys. 18, 489–522. doi: 10.1146 / annurev.es.18.110187.002421
CrossRef Full Text | Google Scholar
Bickford, D., Lohman, D. J., Sodhi, N. S, Ng, P. K., Meier, R., Winker, K., et al. (2007). Kryptiset lajit ikkunana monimuotoisuuteen ja suojeluun. Suuntaukset Ecol. Evol. 22, 148–155. doi: 10.1016/J. puu.2006.11.004
PubMed Abstract | CrossRef Full Text/Google Scholar
Blom, M. P. K., Bragg, J. G., Potter, S., and Moritz, C. (2016). Accounting for uncertainty in gene tree estimation: summary-coalescent species tree inference in a challenging radiation of Australian lizards. Syst. Biol. 66, 352–366. doi: 10.1093/sysbio/syw089
PubMed Abstract | CrossRef Full Text/Google Scholar
Broadhurst, L., Breed, M., Lowe, A., Bragg, J., Catullo, R., Coates, D. J., et al. (2017). Australian kasviston geneettinen monimuotoisuus ja rakenne. Sukeltaja. Ahdistunut. 23, 41–52. doi: 10.1111 / ddi.12505
CrossRef Full Text / Google Scholar
Byrne, M., Coates, D. J., Forest, F., Hopper, S. D., Krauss, S. L., Sniderman, J. M. K., et al. (2014). ”A diverse flora-species and genetic relationships,” in Plant Life on the Sandplains in Southwest Australia, a Global Biodiversity Hotspot, editors H. Lambers (Crawley, WA: University of Western Australia Publishing), 81-99.
PubMed Abstract
Byrne, M., Macdonald, B., and Coates, D. J. (2002). Phylogeographic kuvioita ja suuri cpdna monimuotoisuus sisällä Acacia acuminata (Leguminosae: Mimosoideae). kompleksi Länsi-Australiassa. J. Evol. Biol. 15, 576–587. doi: 10.1046/j.1420-9101.2002.00429.x
CrossRef Full Text/Google Scholar
Carstens, B. C., Pelletier, T. A., Reid, N. M., and Satler, J. D. (2013). Miten epäonnistua lajien karsinnassa. Mol. Ecol. 22, 4369–4383. doi: 10.1111 / mec.12413
PubMed Abstract | CrossRef Full Text/Google Scholar
Coates, D. J. (1988). Geneettinen monimuotoisuus ja populaatiogeenin rakenne harvinaisessa Chittering ruoho wattle, Acacia anomala Court. Aust. J. Bot. 36:273–286. doi: 10.1071/BT9880273
CrossRef Full Text | Google Scholar
Coates, D. J. (2000). Defining conservation units in a rich and fragmented flora: implications for management of genetic resources and evolutionary processes in south-west Australia plants. Aust. J. Bot. 48, 329–339. doi: 10.1071/BT99018
CrossRef Full Text | Google Scholar
Coates, D. J., McArthur, S. L., and Byrne, M. (2015). Merkittävä geneettisen monimuotoisuuden häviäminen, joka johtuu patogeenien aiheuttamasta populaation häviämisestä harvinaisessa endeemisessä Banksia brownii-lajissa (Proteaceae). Biol. Conserv. 192, 353–360. doi: 10.1016 / J.biocon.2015.10.013
CrossRef Full Text/Google Scholar
Colgan, D. J., and Paxton, J. R. (1997). Biokemiallinen genetiikka ja yhteisen gemfishin, Rexea solandri (Scombroidea: Gempylidae), läntisen kannan tunnistaminen., Australia. Maaliskuuta. Freshw. Res. 48, 103-118. doi: 10.1071/MF96048
CrossRef Full Text | Google Scholar
biologista monimuotoisuutta koskeva yleissopimus (2007). Saatavilla verkossa osoitteessa https://www.cbd.int/doc/meetings/cop-bureau/cop-bur-2007/cop-bur-2007-10-14-en.pdf
Cook, C. N., and sgrò, C. M. (2017). Tieteen ja politiikan yhdenmukaistaminen evolutionaarisesti valistuneen suojelun saavuttamiseksi. Conserv. Biol. 31, 501–512. doi: 10.1111 / cobi.12863
PubMed Abstract | CrossRef Full Text/Google Scholar
Cook, L. G., Edwards, R. D., Crisp, M. D., and Hardy, N. B. (2010). Tarvitaanko morfologiaa aina uusiin lajikuvauksiin? Käänteinen. Sys. 24, 322–332. doi: 10.1071/IS10011
CrossRef Full Text | Google Scholar
Cracraft, J. (1983). Lajikäsitteet ja lajiutumisanalyysi. Kurr. Ornitholia. 1, 159–187. doi: 10.1007/978-1-4615-6781-3_6
CrossRef Full Text/Google Scholar
Crandall, K. A., Bininda-Emonds, O. R., Mace, G. M., and Wayne, R. K. (2000). Evoluutioprosessien huomioon ottaminen luonnonsuojelubiologiassa. Suuntaukset Ecol. Evol. 15, 290–295. doi: 10.1016/S0169-5347(00)01876-0
PubMed Abstract | CrossRef Full Text | Google Scholar
Cutter, A. D., and Payseur, B. A. (2013). Genomiset allekirjoitukset valinnasta linkitetyissä kohteissa: lajien välisten erojen yhtenäistäminen. Nat. Pastori Genet. 14, 262–274. doi: 10.1038/nrg3425
PubMed Abstract | CrossRef Full Text/Google Scholar
Dayrat, B. (2005). Kohti integratiivista taksonomiaa. Biol. J. Linn. Soc. 85, 407–415. doi: 10.1111/j.1095-8312.2005.00503.x5
CrossRef Full Text / Google Scholar
de Queiroz, K. (1998). ”The general lineage concept of species, species criteria, and the process of speciation,” teoksessa Forms: Species and Speciation, D. J. Howard, R S. H. Berlocher (New York, NY: Oxford University Press), 57-75.
Google Scholar
de Queiroz, K. (2007). Lajikäsitteet ja lajirajaus. Syst. Biol. 56, 879–886. doi: 10.1080/10635150701701083
PubMed Abstract | CrossRef Full Text | Google Scholar
Degnan, J. H., and Rosenberg, N. A. (2009). Geenipuun ristiriita, fylogeneettinen päättely ja monilajinen yhteenliittymä. Suuntaukset Ecol. Evol. 24, 332–340. doi: 10.1016/J. puu.2009.01.009
PubMed Abstract | CrossRef Full Text/Google Scholar
Devey, D. S., Bateman, R. M., Fay, M. F., and Hawkins, J. A. (2008). Ystäviä vai sukulaisia? Fylogenetiikka ja lajien rajaus kiistellyssä eurooppalaisessa orkideasuvussa Ophrys. Ann. Botti. 101, 385–402. doi: 10.1093/aob/mcm299
PubMed Abstract | CrossRef Full Text | Google Scholar
Dizon, A. E., Lockyer, C, Perrin, W. F., Demaster, D. P., and Sisson, J. (1992). Varastokäsitteen uudelleenarviointi – fylogeografinen lähestymistapa. Conserv. Biol. 6, 24–36. doi: 10.1046 / j.1523-1739.1992. 610024.x
CrossRef Full Text | Google Scholar
Doughty, P., Bourke, G., Tedeschi, L., Pratt, R., Oliver, P., Palmer, R., et al. (2018). Lajirajaus gehyra nana-heimossa (Squamata: Gekkonidae). kompleksi:kryptinen ja poikkeava morfologinen evoluutio Australian Monsoonaalisessa tropiikissa, jossa on neljä uutta lajia. Zootaxa, 4403: 201-244. doi: 10.11646 / zootaxa.4403.2.1
PubMed Abstract | CrossRef Full Text/Google Scholar
Dynesius, M., and Jansson, R. (2014). Lajinsisäisten sukulinjojen pysyvyys: lajiutumisnopeuden laiminlyöty valvonta. Evolution 68, 923-934. doi: 10.1111 / evo.12316
PubMed Abstract / CrossRef Full Text/Google Scholar
Edwards, S. V., Potter, S., Schmitt, C. J., Bragg, J. G., and Moritz, C. (2016). Retikulaatio, divergenssi ja fylogeografia-fylogenetiikka jatkumo. Proc. Natl. Acad. Sci. USA 113, 8025-8032. doi: 10.1073 / pnas.1601066113
CrossRef Full Text/Google Scholar
Faith, D. P. (1992). Conservation evaluation and fylogenetic diversity. Biol. Cons. 61, 1–10. doi: 10.1016/0006-3207(92)91201-3
CrossRef Full Text/Google Scholar
Finston, T. L., Johnson, M. S., Humphreys, W. F., Eberhard, S. M., and Halse, S. A. (2007). Kryptinen lajiutuminen kahteen laajalle levinneeseen maanalaiseen amfipodisukuun heijastaa historiallisia salaojituskuvioita muinaisessa maisemassa. Mol. Ecol. 16, 355–365. doi: 10.1111 / j.1365-294x.2006.03123.x
PubMed Abstract | CrossRef Full Text/Google Scholar
Firestone, K. B., Elphinstone, M. S., Sherwin, W. B., and Houlden, B. A. (1999). Dasyurus maculatus (Dasyuridae: Marsupalia) on uhanalainen lihansyöjä pussieläin. Mol. Ecol. 8, 1613–1625. doi: 10.1046 / j.1365-294x. 1999. 00745.x
CrossRef Full Text/Google Scholar
Fišer, C., Robinson, C. T., and Malard, F. (2018). Kryptiset lajit ikkunana lajikäsityksen paradigman muutokseen. Mol. Ecol. 27, 613–635. doi: 10.1111 / mec.14486
PubMed Abstract | CrossRef Full Text/Google Scholar
Frankham, R. (2010). Geneettisten lähestymistapojen haasteet ja mahdollisuudet biologiseen suojeluun. Biol. Conserv. 143,1919–1927. doi: 10.1016 / J.biocon.2010.05.011
CrossRef Full Text | Google Scholar
Frankham, R., Ballou, J. D., Dudash, M. R., Eldridge, M. D., Fenster, C. B., Lacy, R. C., et al. (2012). Eri lajikäsitteiden vaikutukset luonnon monimuotoisuuden säilyttämiseen. Biol. Conserv. 153, 25–31. doi: 10.1016 / J.biocon.2012.04.034
CrossRef Full Text/Google Scholar
Fraser, D. J., and Bernatchez, L. (2001). Adaptiivisen evoluution säilyttäminen: towards a unified käsite määritellään säilytysyksiköt. Mol. Ecol. 10, 2741–2752. doi: 10.1046/j.1365-294x.2001.t01-1-01411.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Funk, W. C., Mckay, J. K., Hohenlohe, P. A., ja Allendorf, F. W. (2012). Genomiikan valjastaminen konservointiyksiköiden rajaamiseen. Suuntaukset Ecol. Evol. 27, 489–496. doi: 10.1016/J. puu.2012.05.012
PubMed Abstract | CrossRef Full Text/Google Scholar
Garnett, S. T., and Christidis, L. (2017). Taksonomian anarkia haittaa konservointia. Luonto 546, 25-27. doi: 10.1038/546025a
PubMed Abstract | CrossRef Full Text | Google Scholar
Gippoliti, S., Cotterill, F. P. D., Zinner, D., and Groves, C. P. (2018). Taksonomisen tehottomuuden vaikutukset Afrikan sorkka-ja kavioeläinten monimuotoisuuden säilyttämiseen: yleiskatsaus. Biol. Rev. 93, 115-130. doi: 10.1111 / brv.12335
PubMed Abstract | CrossRef Full Text/Google Scholar
Grant, V. (1981). Kasvien Lajiutuminen. New York, NY: Columbia University Press.
PubMed Abstract/Google Scholar
Hamilton, C. W., and Reichard, S. H. (1992). Nykyinen käytäntö alalajien, lajikkeiden ja forma: n käytössä luonnonvaraisten kasvien luokittelussa. Taksoni 41, 485-498. doi: 10.2307 / 1222819
CrossRef Full Text | Google Scholar
Harvey, M. S., Rix, M. G., Framenau, V. W., Hamilton, Z. R., Johnson, ms, Teale, RJ, et al. (2011). Viattomien suojelu: lyhyen kantaman endeemisten taksonien tutkiminen parantaa suojelun tuloksia. Käänteinen. Syst. 25, 1–10. doi: 10.1071/IS11011
CrossRef Full Text | Google Scholar
Hey, J., Waples, R. S., Arnold, M. L., Butlin, R. K., and Harrison, R. G. (2003). Lajien epävarmuuden ymmärtäminen ja kohtaaminen biologiassa ja suojelussa. Suuntaukset Ecol. Evol. 18, 597–603. doi: 10.1016/J. puu.2003.08.014
CrossRef Full Text / Google Scholar
Hoban, S. M., Hauffe, H. C., Perez-Espona, S., Arntzen, J. W., Bertorelle, G., Bryjä, J., et al. (2013). Geneettisen monimuotoisuuden tuominen suojelupolitiikan ja hoidon eturintamaan. Conserv. Genet. Resour. 5, 593–598. doi: 10.1007 / s12686-013-9859-y
CrossRef Full Text/Google Scholar
Isaac, N. J., Mallet, J., and Mace, G. M. (2004). Taksonominen inflaatio: sen vaikutus makroekologiaan ja säilyttämiseen. Suuntaukset Ecol. Evol. 19, 464–469. doi: 10.1016/J. puu.2004.06.004
PubMed Abstract / CrossRef Full Text / Google Scholar
Isaac, N. J., Turvey, S. T., Collen, B., Waterman, C., and Baillie, J. E. (2007). Nisäkkäät reunalla: uhanalaisuuteen ja fylogeniaan perustuvat suojelutavoitteet. PLoS yksi 2: e296. doi: 10.1371 / lehti.pone.0000296
PubMed Abstract | CrossRef Full Text/Google Scholar
Jones, M. R., and Good, J. M. (2016). Kohdennettu talteenotto evolutiivisessa ja ekologisessa genomiikassa. Mol. Ecol. 25, 185–202. doi: 10.1111 / mec.13304
PubMed Abstract | CrossRef Full Text/Google Scholar
Jorger, K. M., and Schrödl, M. (2013). Miten kuvata kryptinen laji? Molekyylitaksonomian käytännön haasteet. Edessä. Zool. 10:59. doi: 10.1186/1742-9994-10-59
PubMed Abstract | CrossRef Full Text/Google Scholar
Laikre, L., Allendorf, F., Aroner, L. C., Baker, S., Gregovich, D. P., Hansen, M. M., et al. (2010). Geneettisen monimuotoisuuden laiminlyönti biologista monimuotoisuutta koskevan yleissopimuksen täytäntöönpanossa. Conserv. Biol. 24, 86–88. doi: 10.1111 / j.1523-1739.2009. 01425.x
PubMed Abstract | CrossRef Full Text/Google Scholar
Laikre, L., Palm, S., and Ryman, N. (2005). Kalojen geneettinen populaatiorakenne: vaikutukset rannikkoalueiden hoitoon. Ambio 34 111–119. doi: 10.1579/0044-7447-34.2.111
PubMed Abstract | CrossRef Full Text/Google Scholar
Maally, T., Laffan, S. W., González-Orozco, C. E., Faith, D. P., Rosauer, D. F., Byrne, M., et al. (2015). Phylodiversity informoimaan conservation policy: a Australian example. Sci. Täydellinen Ympäristö. 534, 131–143. doi: 10.1016 / j.scitotenv.2015.04.113
PubMed Abstract | CrossRef Full Text/Google Scholar
Leache, A. D., and Oaks, J. R. (2017). Yhden nukleotidin polymorfismin (SNP) hyödyllisyys. tiedot fylogenetiikassa. Ann. Rev. Ecol. Syst. 48, 69–84. doi: 10.1146/annurev-ecolsys-110316-022645
CrossRef Full Text | Google Scholar
Llorens, T. M., Macdonald, B., McArthur, S., Coates, D. J., and Byrne, M. (2015). Disjunct, highly divergent genetic lineages within two rare Eremophila (Scrophulariaceae: Myoporeae). species in a biodiversity hotspot: implications for taxonomy and conservation. Biol. J. Linn. Soc. 177, 96–111. doi: 10.1111/boj.12228
CrossRef Full Text | Google Scholar
Love Stowell, S. M., Pinzone, C. A., and Martin, A. P. (2017). Geneettistä monimuotoisuutta koskevien aktiivisten toimien esteiden poistaminen. Biodivers. Conserv. 26, 1753–1765. doi: 10.1007 / s10531-017-1330-z
CrossRef Full Text/Google Scholar
Mace, G. M. (2004). Taksonomian rooli lajien suojelussa. Filos. Trans. R. Soc. B: Biol. Sci. 359, 711–719. doi: 10.1098 / rstb.2003.1454
PubMed Abstract | CrossRef Full Text/Google Scholar
Mallet, J. (2007). Hybridilajittelua. Nature 446, 279-283. doi: 10.1038/nature05706
PubMed Abstract | CrossRef Full Text/Google Scholar
Mayr, E. (1942). Systematiikka ja Lajien synty. New York, NY: Columbia University Press.
Google Scholar
Mee, J. A., Bernatchez, L., Reist, J. D., Rogers, S. M., and Taylor, E. B. (2015). Yksilöidään yksilöitävissä olevat yksiköt erityistä suojelun priorisointia varten: hierarkkinen lähestymistapa, jota sovelletaan järven siika-lajikompleksiin (Coregonus spp.). Evol. Appl. 8, 423–441. doi: 10.1111 / eva.12247
PubMed Abstract | CrossRef Full Text | Google Scholar
Millar, M. A., and Byrne, M. (2013). Cryptic divergent lineages of Pultenaea pauciflora M.B. Scott (Fabaceae: Mirbelieae) exhibit different evolutionary history. Biol. J. Linn. Soc. 108, 871–881. doi: 10.1111/bij.12002
CrossRef Full Text | Google Scholar
Moritz, C. (1994). Defining evolutionarily significant units for conservation. Trends Ecol. Evol. 9, 373–375. doi: 10.1016/0169-5347(94)90057-4
CrossRef Full Text | PubMed Abstract
Moritz, C. (1999). Conservation units and translocations: strategies for Conservation evolutionary processes. Hereditas 130, 217-228. doi: 10.1111 / j.1601-5223.1999. 00217.x
CrossRef Full Text / Google Scholar
Moritz, C. (2002). Strategiat biologisen monimuotoisuuden ja sitä ylläpitävien evoluutioprosessien suojelemiseksi. Syst. Biol. 51, 238–254. doi: 10.1080/10635150252899752
PubMed Abstract | CrossRef Full Text | Google Scholar
Moritz, C., and Potter, S. (2013). Evolutiivisen näkökulman merkitys suojelupolitiikan suunnittelussa. Mol. Ecol. 22, 5969–5971. doi: 10.1111 / mec.12565
PubMed Abstract | CrossRef Full Text | Google Scholar
Moritz, C., Pratt, R. C., Bank, S., Bourke, G., Bragg, J. G., Doughty, P., et al. (2018). Kryptinen sukulinjojen monimuotoisuus, kehon koon eroavuus, ja sympatry lajikompleksissa Australian liskoja (Gehyra). Evolution 27, 54-66. doi: 10.1111 / evo.13380
CrossRef Full Text/Google Scholar
Morrison, W. R., Lohr, J. L., Duchen, P., Wilches, R., Trujillo, D., Mair, M., et al. (2009). Taksonomisen muutoksen vaikutus suojeluun: Tappaako se, pelastaako se vai onko se vain epäolennaista? Biol. Cons. 142, 3201–3206. doi: 10.1016 / J.biocon.2009.07.019
CrossRef Full Text/Google Scholar
Murphy, N. P., Adams, M., Guzik, M. T., and Austin, A. D. (2013). Australian aavikon kevätamphipodeissa on poikkeuksellista mikro-endeemisyyttä. Mol. Fylogeneettiä. Evol. 66, 645–653. doi: 10.1016 / j.ympev.2012.10.013
PubMed Abstract | CrossRef Full Text/Google Scholar
Murphy, N. P., King, R. A., and Delean, S. (2015). Lajit, ESU: t vai populaatiot? Rajaamalla ja kuvaamalla morfologisesti arvoituksellista monimuotoisuutta Australian aavikon kevätamfipodeilla. Käänteinen. Syst. 29, 457–467. doi: 10.1071 / IS14036
CrossRef Full Text/Google Scholar
Nater, A., Mattle-Greminger, M. P., Nurcahyo, A., Nowak, M. G., de Manual, M., Desai, T., et al. (2017). Morfometriset, käyttäytymistieteelliset ja genomiset todisteet uudesta orankilajista. Kurr. Cell Biol. 27, 3487–3498. doi: 10.1016/J. cub.2017.09.047
CrossRef Full Text | Google Scholar
Nicotra, A. B., Chong, C., Bragg, J. G., Ong, C. R., Aitken, N. C., Chuah, A., et al. (2016). Populaatio ja fylogenominen hajoaminen genotyypin mukaan sekvensoimalla Australian Pelargoniumissa. Mol. Ecol. 25, 2000–2014. doi: 10.1111 / mec.13584
PubMed Abstract | CrossRef Full Text/Google Scholar
Nistelberger, H., Byrne, M., Coates, D., and Roberts, J. D. (2014a). Vahva fylogeografinen rakenne tuhatjalkaisessa viittaa Pleistoseeniseen vikarianssiin populaatioiden välillä raudoitetuilla rautamuodostelmilla puolikuivassa Australiassa. PLoS yksi 9: e0093038. doi: 10.1371 / lehti.pone.0093038
PubMed Abstract | CrossRef Full Text/Google Scholar
Nistelberger, H., Gibson, N., Macdonald, B., Tapper, S.-L., and Byrne, M. (2014b). Fylogeografiset todisteet kahdesta mesisestä refugiasta biodiversiteettipesäkkeessä. Perinnöllisyys 113, 454-463. doi: 10.1038 / hdy.2014.46
PubMed Abstract | CrossRef Full Text/Google Scholar
Ogilvie, H. A., Heled, J., Xie, D., and Drummond, A. J. (2016). Laskennallinen suorituskyky ja tilastollinen tarkkuus peto ja vertailuja muihin menetelmiin. Syst. Biol. 65, 381–396. doi: 10.1093/sysbio/syv118
PubMed Abstract | CrossRef Full Text/Google Scholar
Oliver, P. M., Keogh, J. S., and Moritx, C. (2015). New approaches to cataloguing and understanding evolutionary diversity: a perspective from Australian herpetology. Aust. J. Zool. 62, 417–430. doi: 10.1071/ZO14091
CrossRef Full Text | Google Scholar
Oliver, P. M., Laver, R. J., Demello Martins, F., Pratt, R. C., Hunjan, S., and Moritz, C. (2017). Selkärankaisten endemismin Uusi hotspot ja evolutiivinen refugium trooppisessa Australiassa. Sukeltaja. Ahdistunut. 23, 53–66. doi: 10.1111 / ddi.12506
CrossRef Full Text/Google Scholar
Oliver, P. M., and Lee, M. S. Y. (2010). Kasvitieteelliset ja eläintieteelliset koodit haittaavat biodiversiteettitutkimusta, koska ne estävät nimeämättömien uusien lajien julkaisemisen. Taksoni 59, 1201-1205. doi: 10.2307/20773990
CrossRef Full Text | Google Scholar
Padial, J., Miralles, A., De la Riva, I., and Vences, M. (2010). Taksonomian integratiivinen tulevaisuus. Edessä. Zool. 7:16. doi: 10.1186/1742-9994-7-16
PubMed Abstract | CrossRef Full Text/Google Scholar
Palsbøll, P. J., Bérubé, M., and Allendorf, F. W. (2007). Hallintoyksiköiden tunnistaminen populaatiogeneettisiä tietoja käyttäen. Suuntaukset Ecol. Evol. 22, 11–16. doi: 10.1016/J. puu.2006.09.003
PubMed Abstract | CrossRef Full Text/Google Scholar
Patton, J. L., and Smith, M. F. (1994). Parafyly, polyfyly, ja luonne lajien rajoja taskurotta (suku Thomomys). Syst. Biol. 43, 11–26. doi: 10.1093/sysbio / 43.1.11
CrossRef Full Text | Google Scholar
Pierson, J. C., Coates, D. J., Oostermeijer, J. G. B., Beissinger, S. R., Bragg, J. G., Sunnucks, P., et al. (2016). Perintötekijät uhanalaisten lajien elvytyssuunnitelmissa kolmella mantereella. Edessä. Ecol. Environ. 14, 433–440. doi: 10.1002 / maksu.1323
CrossRef Full Text/Google Scholar
Potter, S., Bragg, J. G., Peter, B. M., Bi, K., and Moritz, C. (2016). Fylogenomics vinkkejä: päättely lineages ja niiden demografinen historia trooppinen lisko, Carlia amax. Mol. Ecol. 25, 1367–1380. doi: 10.1111 / mec.13546
PubMed Abstract | CrossRef Full Text/Google Scholar
Potter, S., Xue, A. T., Bragg, J. G., Rosauer, D. F., Roycroft, E. J., and Moritz, C. (2018). Pleistoseenin ilmastomuutokset ajavat monipuolistumaan trooppisella savannilla. Mol. Ecol. 27, 520–532. doi: 10.1111 / mec.14441 / full
PubMed Abstract / CrossRef Full Text/Google Scholar
Rabosky, D. L., Hutchinson, M. N., Donnellan, S. C., Talaba, A. L., and Lovette, I. J. (2014). Lajirajojen fylogeneettinen purkaminen laajalle levinneessä Australiankinkkuryhmässä (Scincidae: Ctenotus). Mol. Fylogeneettiä. Evol. 77, 71–82. doi: 10.1016 / j.ympev.2014.03.026
PubMed Abstract | CrossRef Full Text/Google Scholar
Ralls, K., Ballou, J. D., Dudash, M. R., Eldridge, M. D. B., Fenster, C. B., Lacy, R. C., et al. (2018). Vaativat paradigman muutosta hajanaisten populaatioiden geneettisessä hallinnassa. Conserv. Lett. 11, 1–6. doi: 10.1111 / con.124126
CrossRef Full Text/Google Scholar
Rannala, B. (2015). Lajirajauksen taidetta ja tiedettä. Kurr. Zool. 61, 846–853. doi: 10.1093/czoolo/61.5.846
CrossRef Full Text | Google Scholar
Riedel, A., Sagata, K., Suhardjono, Y. R., Tänzler, R., and Balke, M. (2013). Integratiivinen taksonomia on the fast track-towards more sustainability in biodiversity research. Edessä. Zool. 10:15. doi: 10.1186/1742-9994-10-15
PubMed Abstract | CrossRef Full Text | Google Scholar
Rieseberg, L. H., and Willis, J. H. (2007). Kasvien lajiutuminen. Tiede 317, 910-914. doi: 10.1126 / tiede.1137729
PubMed Abstract | CrossRef Full Text/Google Scholar
Rosauer, D. F., Blom, M. P. K., Bourke, G., Catalano, S., Donnellan, S., Mulder, E., et al. (2016). Fylogeography, hotspots ja suojelu painopisteet: esimerkki yläpäässä Australiassa. Biol. Cons. 204, 83–93. doi: 10.1016 / J.biocon.2016.002
CrossRef Full Text | Google Scholar
Rosauer, D. F., Byrne, M., Blom, M. P. K., Coates, D. J., Donnellan, S., Doughty, P., et al. (2018). Australian Kimberleyssä tapahtuva evolutiivisen monimuotoisuuden suojeluun tähtäävä real-world conservation planning syrjäyttää epävarman taksonomian Conserv: n. Lett. 11: e12438. doi: 10.1111 / con.12438
CrossRef Full Text/Google Scholar
Rosenblum, E. B., Sarver, B. A. J., Brown, J. W., Des Roches, S., Hardwick, K. M., Hether, T. D., et al. (2012). Kultakutri täyttää Santa Rosalian: lyhytaikainen lajiutumismalli selittää eriytymisen eri aikaväleillä. Evol. Biol. 39, 255–261. doi: 10.1007 / s11692-012-9171-x
PubMed Abstract | CrossRef Full Text/Google Scholar
Roux, C., Fraïsse, C., Romiguier, J., Anciaux, Y., Galtier, N., and Bierne, N. (2016). Se valaisee lajiutumisen harmaata vyöhykettä genomierojen jatkumossa. PLoS Biol. 14: e2000234. doi: 10.1371 / lehti.pbio.2000234
PubMed Abstract | CrossRef Full Text/Google Scholar
Ryder, O. A. (1986). Lajien suojelu ja systematiikka-alalajien dilemma. Suuntaukset Ecol. Evol. 1, 9–10. doi: 10.1016/0169-5347 (86) 90059-5
CrossRef Full Text | Google Scholar
Sampson, J. F., Byrne, M., Yates, C. J., Gibson, N., Thavornkanlapachai, R., Stankowski, S., et al. (2014). Nykyinen siitepölyvälitteinen geenimaahanmuutto kuvastaa harvinaisen pensaan historiallista eristyneisyyttä hajanaisessa maisemassa. Perinnöllisyys 112, 172-181. doi: 10.1038 / hdy.2013.89
PubMed Abstract | CrossRef Full Text/Google Scholar
Santamaría, L., and Méndez, P. F. (2012). Biologista monimuotoisuutta koskevan politiikan kehitys: nykyiset puutteet ja tulevat tarpeet. Evol. Appl. 5, 202–218. doi: 10.1111 / j.1752-4571.2011.00229.x
PubMed Abstract | CrossRef Full Text/Google Scholar
Schlick-Steiner, B. C., Steiner, F. M., Seifert, B., Stauffer, C., Christian, E., and Crozier, R. H. (2010). Integratiivinen taksonomia: monilähteinen lähestymistapa biologisen monimuotoisuuden tutkimiseen. Annu. Pastori Entomol. 55, 421–438. doi: 10.1146/annurev-ento-112408-085432
PubMed Abstract | CrossRef Full Text | Google Scholar
Seehausen, O, Takimoto, G., Roy, D., and Jokela, J. (2008). Lajiutumisen kääntyminen ja biodiversiteetin dynamiikka hybridisaation kanssa muuttuvissa ympäristöissä. Mol Ecol. 17, 30–44. doi: 10.1111 / j.1365-294x.2007.03529.x
PubMed Abstract | CrossRef Full Text/Google Scholar
Shepherd, K. A., Thiele, K. R., Sampson, J., Coates, D. J., and Byrne, M. (2015). Harvinainen, uusi laji Atriplex (Chenopodiaceae), joka käsittää kaksi geneettisesti erillistä mutta morfologisesti arvoituksellista populaatiota kuivissa Länsi-Australiassa: implications for taxonomy and conservation. Aust. Syst. Botti. 28, 234–245. doi: 10.1071/Sb15029
CrossRef Full Text/Google Scholar
Singhal, S., and Bi, K. (2017). Historia siivoaa sotkut: ajan vaikutus ajamiseen divergenssi ja introgressio trooppisella ompeluvyöhykkeellä. Evolution 71, 1888-1899. doi: 10.1111 / evo.13278
PubMed Abstract | CrossRef Full Text/Google Scholar
Singhal, S., Hoskin, C., Couper, P., Potter, S., and Moritz, C. (2018). Framework for resolving cryptic species: a case study from the lizards of the Australian wet tropics. Syst. Biol. . doi: 10.1093/sysbio/syy
PubMed Abstract | CrossRef Full Text/Google Scholar
Singhal, S., and Moritz, C. (2013). Lisääntymis eristäminen välillä fylogeographic lineages asteikot eroavaisuuksia. Proc. Roy. Soc B 280: 20132246. doi: 10.1098 / rspb.2013.2246026
PubMed Abstract | CrossRef Full Text/Google Scholar
Steane, D. A., Potts, B. M., McLean, E., Collins, L., Prober, S. M., Stock, W. D., et al. (2015). Genomin laajuiset luotaukset paljastavat kryptisen väestörakenteen kuivaksi sopeutuneessa eukalyptuksessa. Tree Genet. Genomes 11: 33. doi: 10.1007 / s11295-015-0864-z
CrossRef Full Text | Google Scholar
Struck, T. H., Feder, J. L., Bendiksby, M., Birkeland, S., Cerca, J., Gusarov, V. I., et al. (2017). Löytää evoluutioprosesseja piilotettuna kryptisiin lajeihin. Suuntaukset Ecol. Evol. 33, 153–163. doi: 10.1016/J. puu.2017.11.007
PubMed Abstract | CrossRef Full Text/Google Scholar
Sukumaran, J., and Knowles, L. L. (2017). Monilajinen yhteenliittymä rajaa rakennetta, ei lajeja. Proc. Nat. Aca. Sci. 114, 1607-1612. doi: 10.1073 / pnas.1607921114
PubMed Abstract | CrossRef Full Text/Google Scholar
Swarts, N. D., Clements, M. A., Bower, C. C., and Miller, J. T. (2014). Määrittelemällä suojeluyksiköt morfologisesti samanlaisten, seksuaalisesti petollisten, erittäin uhanalaisten orkideoiden kompleksissa. Biol. Cons. 174, 55–64. doi: 10.1016 / J.biocon.2014.03.017
CrossRef Full Text/Google Scholar
Tautz, D., Arctander, P., Minelli, A., Thomas, R. H., and Vogler, A. P. (2003). Vetoomus DNA: n taksonomian puolesta. Suuntaukset Ecol. Evol. 18, 70–74. doi: 10.1016/S0169-5347(02)00041-1
CrossRef Full Text/Google Scholar
Taylor, H. R., Dussex, N., and van Heezik, Y. (2017). Suojeluun liittyvän genetiikan kuilun kurominen umpeen tunnistamalla suojelun harjoittajien toteuttamiselle esteitä. Glob. Ecol. Conserv. 10, 231–242. doi: 10.1016 / j.gecco.2017.04.001
CrossRef Full Text / Google Scholar
Thomson, S. A., Pyle, R. L., Ahyong, S. T., Alonso-Zarazaga, M., Ammirati, J., Araya, J. F., et al. (2018). Tieteeseen perustuva taksonomia on välttämätöntä maailmanlaajuiselle suojelulle. PLoS Biol. 16: e2005075. doi: 10.1371 / lehti.pbio.2005075
PubMed Abstract | CrossRef Full Text/Google Scholar
Vieites, D. R., Wollenberg, K. C., Andreonec, F., Köhler, J., Glaw, F., and Vences, M. (2009). Madagaskarin biologisen monimuotoisuuden laaja aliarviointi, josta on osoituksena integratiivinen sammakkoeläinten inventointi. Proc. Natl. Acad. Sci. USA 106, 8267-8272. doi: 10.1073 / pnas.0810821106
CrossRef koko teksti
Vogler, A. P., and de Salle, R. (1994). Suojeluyksiköiden diagnosointi. Cons. Biol. 8, 354–363. doi: 10.1046 / j.1523-1739.1994. 08020354.x
CrossRef Full Text/Google Scholar
Waples, R. S. (1991). Tyynenmerenlohi, Oncorhynchus spp., ja Uhanalaisuuslain mukainen ”lajin” määritelmä. Maaliskuuta. Kala. Rev. 53, 11-22.
Google Scholar
Waples, R. S. (1995). ”Evolutionarily significant units and the conservation of biological diversity under the Endangered Species Act” teoksessa Evolution and The Aquatic Ecosystem: Defining Unique Units in Population Conservation, eds J. L. Nielsen (Bethesda, MD:American Fisheries Society), 8-27.
Google Scholar
Waples, R. S., Gustafson, R. G., Weitkamp, L. A., Myers, J. M., Jjonson, O. W., Busby, P. J., et al. (2001). Luonnehtivat Tyynenmeren luoteisosan lohien monimuotoisuutta. J. Fish Biol. 59, 1–41. doi: 10.1111 / j.1095-8649.2001.tb01376.x
CrossRef Full Text/Google Scholar
Waples, R. S., Nammack, M., Cochrane, J. F., and Hutchings, J. A. (2013). A tale of two acts: endangered species listing practices in Canada and the United States. BioScience 63, 723-734. doi: 10.1093 / bioscience / 63.9.723
CrossRef Full Text/Google Scholar
Waples, R. S., Punt, A. E., and Cope, J. M. (2008). Geenitiedon integrointi meren luonnonvarojen hallintaan: miten voimme tehdä sen paremmin? Kala 9, 423-449. doi: 10.1111/j.1467-2979.2008.00303.x
CrossRef Full Text/Google Scholar
Wege, J. A., Thiele, K. R., Shepherd, K. A., Butcher, R., Macfarlane, T. D., and Coates, D. J. (2015). Strateginen taksonomia biodiversiteettimaisemassa: uudenlainen lähestymistapa harvinaisten ja huonosti tunnettujen kasvistojen suojelutulosten maksimointiin. Biodivers. Conserv. 24, 17–32. doi: 10.1007 / s10531-014-0785-4
CrossRef Full Text/Google Scholar
Whitlock, R. (2014). Relations between adaptive and neutral genetic diversity and ecological structure and functioning: a meta-analysis. J. Ecol. 102, 857–872. doi: 10.1111/1365-2745.12240
PubMed Abstract | CrossRef Full Text | Google Scholar
Wiley, E. O. (1978). Evolutionaarinen lajikäsitys harkittiin uudelleen. Syst. Biol. 27, 17–26. doi: 10.2307/2412809
CrossRef Full Text | Google Scholar
Wilson, E. O. (2017). Biodiversiteettitutkimus vaatii enemmän saappaat jalassa. Nat. Ecol. Evol. 1, 1590–1591. doi: 10.1038 / s41559-017-0360-y
PubMed Abstract | CrossRef Full Text/Google Scholar
Yang, Z. H., and Rannala, B. (2014). Ohjaamaton lajien rajaus käyttäen DNA-sekvenssitietoja useista lokuksista. Mol. Biol. Evol. 31, 3125–3135. doi: 10.1093/molbev/msu279
PubMed Abstract | CrossRef Full Text | Google Scholar
Yeates, D. K., Seago, A., Nelson, L., Cameron, S. L., Joseph, L., and Trueman, W. H. (2011). Integratiivinen taksonomia vai iteratiivinen taksonomia? Sys. Ent. 36, 209–217. doi: 10.1111/j.1365-3113.2010.00558.x
CrossRef Full Text | Google Scholar