XIX vuosisadan alussa von Reckingausen havaitsi, että verisuonia peitti solulevy. Tämä yksisoluinen paksu kerros koostuu endoteelisoluista, jotka reunustavat veren ja imusuonten sisäpintaa. Ihmisillä endoteelikerroksen kokonaispinta-ala on arviolta noin 35 m2 ja endoteelisoluja on noin 1-1, 6 103. Valtimoiden, laskimoiden ja hiussuonien yhteispituus on noin 90000 km (enemmän kuin matkustaminen kahdesti maailman ympäri). Endoteelisolut ovat litteitä ja liittyneet toisiinsa soluliitosten avulla. Niiden päätehtävä on välittäjinä veren ja muiden kudosten välillä ja edistää veren ominaisuuksia sekä normaaleissa että patologisissa kudoksissa. Ne ovat myös tärkeitä veren ja kudosten immuunisolujen vaihdossa.
1. Morfologia
endoteelisolut ovat hyvin litistyneitä soluja niin paljon, että niiden tuma on solun pitempi rakenne, vaikka Tuma olisi myös litistynyt. Solumuoto mukautuu kanavaan, jossa endoteelisolu on vuorattu (Kuva 1, 2 ja 3). Kapeammissa hiussuonissa endoteelisolut voivat laajentaa sytoplasmaansa koko verisuonen kehälle niin, että kanavassa on rivi endoteelisoluja. Suuremmissa kanavissa, kuten valtimoissa ja laskimoissa, tarvitaan monia endoteelisoluja kanavan koko kehän peittämiseen.
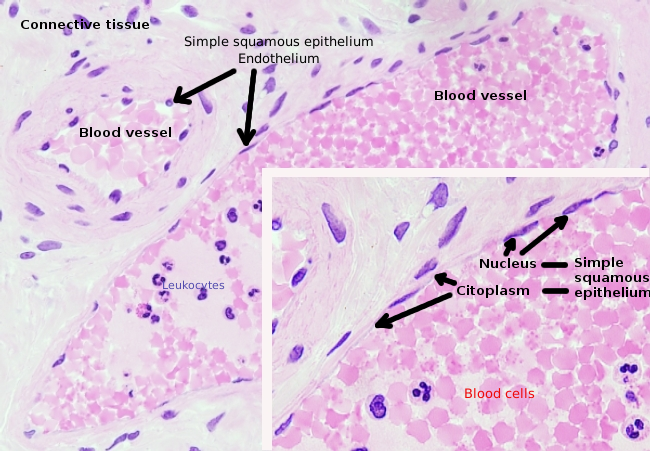
Perinukleaarinen sytoplasma sisältää eniten mitokondrioita ja muita organelleja, kuten Golgin laitteen, kun taas perifeerinen sytoplasma on ohuempaa ja sillä on vähän organelleja, joskin endoplasmainen retikulum löytyy läheltä plasmakalvoa. Endoteelisoluissa on monia vesikkeleitä, joista useimmat ovat endosyyttisiä vesikkeleitä.
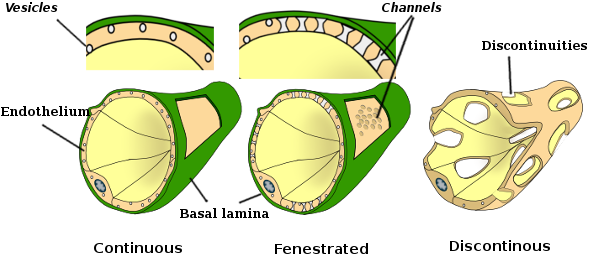
joillakin alueilla, kuten maksassa, munuaiskuoressa ja umpirauhasissa, hiussuonien endoteelisoluissa on huokosia tai pieniä kohtia, joita kutsutaan fenestratrioneiksi. Ne ovat noin 60-70 nm: n läpimittaisia reittejä, joiden kapea käytävä on noin 5 nm ja jotka kommunikoivat suoraan veren ja imun muiden kudosten kanssa. Hiussuonia, joilla on tämäntyyppisiä endoteelisoluja, kutsutaan fenestroituneiksi hiussuoniksi. Huokoset jakautuvat ryhmiin ja niiden tiheys riippuu endoteelin tyypistä.
endoteelisolut erotetaan ympäröivästä kudoksesta solunulkoisella matriisikerroksella, jota kutsutaan tyvilaminaksi. Endoteelisolu osallistuu tähän kerrokseen proteiineilla laminiini, fibronektiini ja kollageenityypit II, IV ja V.
lähde ja proliferaatio
Vaskulogeneesi tarkoittaa uusien endoteelisolujen muodostumista kudoksissa, joissa ei ole ennestään verisuonia. Se tapahtuu alkiossa. Aikuisilla ja myös alkioilla syntyy uusia verisuonia ja siten uusia endoteelisoluja jo nykyisistä verisuonista. Tätä prosessia kutsutaan angiogeneesiksi. Uusia endoteelisoluja tuotetaan haaroittamalla tai jakamalla veri-ja imusuonia. Angiogeneesi on yleistä kasvavissa kudoksissa tai raskaan remodeling, kuten kohtu nisäkkäiden naaraat, tai sairaudet, kuten kasvaimet, tulehdus ja haavat.
Vaskulogeneesi
nisäkkäiden ensimmäiset endoteelisolut syntyvät gastrulaation jälkeen alkionkehityksen aikana. Ne erilaistuvat angioblasteiksi kutsutuista soluista, muodostavat ryhmiä ja järjestäytyvät lyhyiksi verisuoniksi. Tämä prosessi tapahtuu ensin alkion vitelin-pussissa. Nämä ensimmäiset verisuonet kasvavat ja kytkeytyvät toisiinsa muodostaen verkoston. Myöhemmin ne värväävät fibroblasteja ja lihassoluja. Tämä alkuperäinen verkko käy läpi jatkuvan uudelleenmuotoutumisen seuraavien kehitysvaiheiden aikana. Alkioissa kaikki endoteelisolut ovat aluksi samanlaisia, mutta ne erilaistuvat myöhemmin suoneen, valtimoon, kapillaariin ja imusuonen endoteeliin. Useat molekyylit, kuten FGF2, BMP4, IHH ja VEGF, indusoivat erillisiä erilaistumisreittejä. Notch inducer näyttää olevan erittäin tärkeä, koska se johtaa valtimon endoteeli, kun taas sen esto johtaa vein endoteeli. Lymfaattinen endoteeli muodostuu kardinaalilaskimon endoteelista ja tarvitsee ilmaisun COUPF-II ja SOX18. Lisäksi endoteeli erilaistuu sen mukaan, missä elimessä se sijaitsee. Esimerkiksi endoteeli muodostaa tiiviisti suljetun kerroksen aivoihin veri-aivoesteen muodostamiseksi, kun taas se fenestrated on löyhästi järjestäytynyt maksassa suosimaan molekyylien vaihtoa veren kanssa.
koska endoteelisolut järjestäytyvät valtimo-ja laskimokanaviin, jotkut endoteelisolut synnyttävät hematopoieettisia soluja erilaistumisprosessilla, joka tunnetaan ENDOTEELIS-hematopoieettisena siirtymänä (EHT). Tämä prosessi tapahtuu myös joissakin aikuisten kudoksissa, kuten istukan ja muiden kudosten muodostumisen aikana raskauden aikana.
angiogeneesi
angiogeneesi on uusien verisuonten ja endoteelin muodostumista haaroittamalla tai jakamalla olemassa olevia verisuonia. Aikuisilla endoteelisolujen proliferaatioaste on hyvin alhainen. Esimerkiksi nisäkkäillä endoteelisolu voi jakautua kerran kuukaudessa tai odottaa useita vuosia. Se tarkoittaa, että angiogeneesi on harvinaista kudoksissa normaalissa fysiologiassa, lukuun ottamatta naisten lisääntymiselimiä. Se voi kuitenkin aktivoitua patologisissa prosesseissa, kuten kudosvaurioissa ja kasvaimissa. Eräät aineet indusoivat angiogeneesiä, kuten endoteelikasvutekijä (EGF), hapan EGF, fibroblastikasvutekijä (FGF), transformoiva kasvutekijä (TGF) ja prostaglandiinit. Angiogeneesi alkaa, kun ympäröivistä kudoksista vapautuu angiogeenisiä aineita. Läheisen verisuonen endoteelisolu, yleensä venulesta, alkaa siirtyä angiogeeniseen signaalilähteeseen vetäen muita endoteelisoluja, jotka alkavat lisääntyä ja muodostaa uuden astian. Nämä endoteelisolut menettävät hetkellisesti tiiviit yhteytensä, mikä johtaa tyvikalvon paljastumiseen, jonka endoteelisoluista vapautuvat entsyymit pilkkovat nopeasti. Jotkut endoteelisolut menevät pilkkoutuneen tyvikalvon läpi samalla kun lisääntyminen jatkuu. Kaikki tämä prosessi mahdollistaa uuden verisuonen venymisen ja haarautumisen, kunnes vaskulogeneesi-signaali katoaa. Verisuonten kärjet sulautuvat muihin verisuoniin muodostaen suljettuja piirejä ja mahdollistaen veren virtauksen ilman kuolleita päätteitä.
dorsaalisesta aortasta ja endokardiumista on löydetty alueita, joissa on endoteelisolujen esiasteita. Mesenkyymisolut voivat erilaistua endokardium-endoteelisoluista ja muodostaa sydämen kolmiliuskaläpän ja joitakin fibroblasteja, mutta eivät kardiomiosyyttejä. Endoteelisolut ovat niin järkeviä ulkoisille signaaleille, että endoteelin eheyden säilyttäminen on aktiivinen prosessi, jota välittävät muut signaalit, kuten FGF. Aktiivinen prosessi ei ole vain eheyden vuoksi, vaan myös solun fenotyypin säilyttämiseksi. Kun endoteelisolut eivät saa kunnollisia signaaleja, ne kuolevat apoptoosiin tai muuttuvat mesenkyymisoluiksi, jotka saattavat syntetisoida runsaasti solunulkoista matriisia. Siirtyminen endoteelista mesenkymaalisoluun on joidenkin sairauksien, kuten arterioskleroosin ja kuitumikardiitin taustalla.
funktio
ajatus siitä, että endoteeli on vain imusuonten ja verisuonten passiivinen vuorikerros, on muutettava. Endoteelin toiminnot ovat monipuolisia ja välttämättömiä eliölle. Itse asiassa se on sydän-ja imukanavien fyysinen rakenne, mutta se säätelee myös molekyylien vaihtoa aluksen lumenin ja ympäröivien kudosten välillä, ja se vaikuttaa veren fysiologisiin ominaisuuksiin. Fyysisen esteen lisäksi endoteelisolut kehittävät eritys -, aineenvaihdunta-ja immuunitoimintoja. Ne muuttavat fysiologista käyttäytymistään, johon vaikuttavat molekyylit, kuten kasvutekijät, koagulantit ja antikoagulantit, low density lipoproteiinit, typpioksidi, serotoniini, enkefaliini ja monet muut. Endoteelisoluissa on reseptoreita kaikille näille aineille.
este
endoteelisolut muodostavat kerroksen, joka yleensä toimii esteenä veren ja sitä ympäröivien kudosten välillä. Endoteelisolujen välinen koheesio täyttyy soluliitoksissa, kuten tiukoissa liitoksissa ja kiinnittyneissä liitoksissa. Myös aukkoliitoksia on havaittu, vaikka niiden tärkein tehtävä on vierekkäisten solujen välinen viestintä. Endoteelisolut voivat moduloida näitä kiinnikkeitä ja muuttaa esteen läpäisevyyttä, mikä voi vaikuttaa paitsi molekyyleihin myös endoteelin ylittäviin soluihin. Joissakin elimissä, kuten maksassa, endoteelisolut ovat kuitenkin paljon löyhemmin pakattuja ja jättävät paljon vapaata tilaa, joten endoteelia tuskin voidaan pitää esteenä.
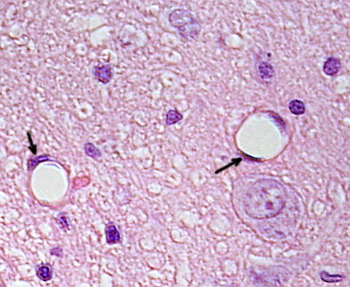
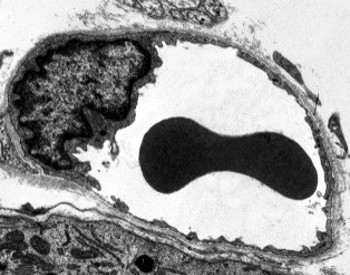
Transmissioelektronimikroskopiakuvissa näkyy endoteelisolujen sytoplasmassa useita vesikkeleitä. Niiden arvellaan osallistuvan solunsisäiseen kuljetukseen apikaalisten (päin verta) ja basolateraalisten kalvojen (päin tyvilaminaa) välillä. Tämä kuljetustyyppi tunnetaan nimellä transkytoosi. On mielenkiintoista, että transkytoosirakkulat ovat runsaampia kapillaarien endoteelisoluissa kuin suurempien verisuonten soluissa. Se osoittaa, että hiussuonien endoteelisoluissa on voimakkaampi molekyylien vaihto veren ja kudosten välillä, kun taas halkaisijaltaan suurissa verisuonissa ne osallistuvat pääasiassa veren johtamiseen. Joissakin endoteelisoluissa, jotka tunnetaan fenestraatteina, on huokosia tai hyvin pieniä kanavia, jotka yhdistävät veren suoraan ympäröiviin kudoksiin, jolloin jotkin pienikokoiset molekyylit voivat ylittää endoteelin joutumatta minkään endoteelisolujen sytoplasmaan (Kuva 4). Lopuksi on elimiä, joissa on hyvin läpäisevä endoteeli. Sinusoidit ovat maksassa verisuonia, joissa endoteelisolut jättävät vapaata tilaa toistensa väliin,eikä esteenä toimivaa toimintoa juuri ole.
monet solutyypit kulkevat veressä kohti kohde-elimiään kehossa. Siellä ne risteävät verisuonten endoteelin, yleensä kapillaarien jälkeisten laskimoiden tasolla. Se tarkoittaa, että endoteelisolujen on muokattava soluliitoksia, jotta solut pääsevät endoteelikerroksen läpi. Leukosyytit poistuvat verisuonesta tunnistamalla ja ankkuroimalla tiettyihin endoteelisolujen apikaalisen kalvon molekyyleihin. Selektiinit, integriinit ja immunoglobuliinit ovat vastuussa leukosyyttien tunnistamisesta ja kiinnittymisestä endoteelikerrokseen. Selektiinit aloittavat leukosyyttien ankkuroinnin, jotka rullaavat endoteelin pinnan yli. Tämä alkuperäinen tartunta on heikko ja palautuva.
monet leukosyytit poistuvat verestä tulehdusprosessien aikana siirtyäkseen sairastuneisiin kudoksiin. Kemokiinit ovat leukosyyttejä puoleensavetäviä molekyylejä, jotka vapautuvat vaurioituneista kudoksista ja liittyvät endoteelisolujen glykokalixiin. Endoteelin pinnalla vierivät leukosyytit, joilla on kemokiineja, aktivoituvat, solukiinnitys voimistuu ja leukosyytti jää paikalleen. Leukosyyttien aktivoituminen johtaa sen integriinien aktivoitumiseen, sillä ne tunnistavat endoteelisolun immunoglobuliinit. Nämä immunoglobuliinit ilmentyvät endoteelisolukalvoissa sen jälkeen, kun kemokiinit ovat aktivoineet solun. Solun ja solun välinen adheesio nostaa endoteelisolujen kalsiumpitoisuutta ja johtaa soluliitosten epäjärjestäytymiseen ja sytoplasman vetäytymiseen. Tällä tavoin leukosyytit voivat siirtyä endoteelisolujen rajalle ja ylittää endoteelin. Adheesiomolekyylit osallistuvat myös tähän leukosyyttien liikkeeseen.
veren ominaisuudet
endoteelifunktiot ovat monimutkaisempia kuin vain endoteelin poikki kulkevien molekyylien ja solujen kontrollointi. Endoteelisolut osallistuvat myös verenpaineeseen, hyytymiseen ja joihinkin muihin veren ominaisuuksiin.
primitiivisen verenkiertojärjestelmän arvellaan syntyneen 600 miljoonaa vuotta sitten selkärangattomissa, mutta siltä puuttui endoteeli. Endoteelisolut ilmestyivät 100 miljoonaa vuotta sitten tarjoten verelle laminaarisemman vuon (ei turbulenttista) ja siten tehokkaamman kaasunvaihdon.
endoteelisolut moduloivat verenpainetta vapauttamalla aineita, jotka vaikuttavat verisuonten sileään lihakseen. Niistä vapautuu typpioksidia (NO) ja prostasykliiniä, jotka rentouttavat verisuonten sileää lihasta. Ne vapauttavat myös endoteliinia ja verihiutaleiden aktivoivaa tekijää, jotka molemmat pienentävät verisuonen halkaisijaa. Typpioksidi vapautuu konstitutiivisesti ja antaa oikean lihaskunnon, estää verihiutaleiden aggregaatiota ja leukosyyttien adheesiota. Endoteliini on vahva vasokonstriktori. Miten endoteelisolut päättävät, mitä molekyyliä on vapautettava, ei ole vielä selvää, mutta apikaalisten kalvojensa mekanoreseptorit, jotka kykenevät tuntemaan veren vuo-ominaisuudet, voivat olla uskottavia.
normaaliolosuhteissa endoteelisoluista vapautuu vereen molekyylejä, jotka auttavat ylläpitämään oikeaa juoksevuutta. Ne toimivat kahdella tasolla: juoksevuus (antikoagulantit) ja verihiutaleiden aggregaation estäminen (antitromboottiset aineet). Proteiinit C ja S ovat tärkeitä veren juoksevuuteen vaikuttavia molekyylejä. S-proteiinin kanssa kompleksin muodostava C-proteiini inaktivoi hyytymistekijät VIIIa ja Va. S-proteiinia syntetisoivat endoteelisolut. Lisäksi endoteeliglykoli sisältää hepariinin kaltaista glykosaminoglykaania, joka pystyy inaktivoimaan trombiinin.
antikoagulanttitoimintaansa liittyen endoteelisolut vapauttavat typpioksidia ja prostasykliiniä, jotka molemmat nostavat verihiutaleiden syklistä AMP-arvoa ja vaikeuttavat verihiutaleiden aggregaatiota. Nämä kaksi molekyyliä vapautuvat jatkuvasti vereen. Endoteelisoluilla on apikaalisissa kalvoissaan ectonukleaaseja, jotka poistavat ATP: tä ja ADP: tä, jotka molemmat edistävät voimakkaasti verihiutaleiden aggregaatiota. Lisäksi endoteelisoluista vapautuu aktivaattorimolekyyli, joka muuttaa plaminogeenin plasmiiniksi, mikä suosii trombien poistamista.
kaikki nämä molekyylireitit voivat muuttua, kun endoteelisolut saavat joitakin signaaleja tai kudos vaurioituu, mikä johtaa veren hyytymiseen ja verihiutaleiden aggregaatioon. Tällöin endoteelisoluista tulee hyytymisen ja tromboosin aktiivisia osallistujia.
immuunipuolustus
endoteelisoluilla on merkittävä rooli immuunipuolustuksessa ja ne osallistuvat kahteen mekanismiin: antigeenien esittämiseen T-lymfosyyteille ja immuunisolujen värväämiseen. Yhdessä makrofagien kanssa endoteelisolut voivat esittää antigeenejä T-lymfosyyteille, koska ne ilmentävät konstitutiivisesti MHC-I: tä (major histocompatibility complex) ja ne voidaan indusoida ilmaisemaan MHC-II: ta, jotka molemmat ovat välttämättömiä antigeenin esittämiseen. Endoteelisolut kykenevät aktivoimaan immuunimuistia, mutta eivät uusia T-lymfosyyttejä. Endoteelisolu-ja T-lymfosyyttien välillä on kaksisuuntainen aktivaatio, jolloin endoteelisolu vapauttaa tulehdussoluja vetäviä molekyylejä ja veren leukosyyttejä ankkuroivia express-adheesiomolekyylejä.
bibliografia
Cines BD, Pollak ES, Buck CA, Loscalzo J, Zimmerman GA, McEver Rp, Pober JS, Wick TM, Konkle BA, Schwartz BS, Barnathan ES, McCrae KR, Hug BA, Schmidt A-M, Stern DM 1998. Endoteelisolut fysiologiassa ja verisuonisairauksien patofysiologiassa. Journal of the american society of hematology. 91:3527-3561 ![]()
Fajardo LF. 1988. Endoteelisolujen monimutkaisuus. American journal of clinical pathology. 92:241-250.
Michiels C. 2003. Endoteelisolu toimii. Journal of cellular physiology. 196: 430-443 ![]()