i begyndelsen af 1960 ‘ erne begyndte biologer at undersøge, hvordan proteiner i forskellige arter udvikler sig på Sekvensniveau (1962, margoliash 1963, Doolittle & blomb lystrack 1964). De analyserede proteiner omfattede hæmoglobin (Sukkerkandl & Pauling 1962), cytochrom C (Margoliash 1963) og fibrinopeptider (Doolittle & Blomb Lutck 1964). Disse tidlige undersøgelser førte til en bemærkelsesværdig opdagelse: det så ud til, at antallet af forskelle mellem proteinsekvenser af forskellige arter var nogenlunde proportional med tiden siden artsdivergens (Figur 1).
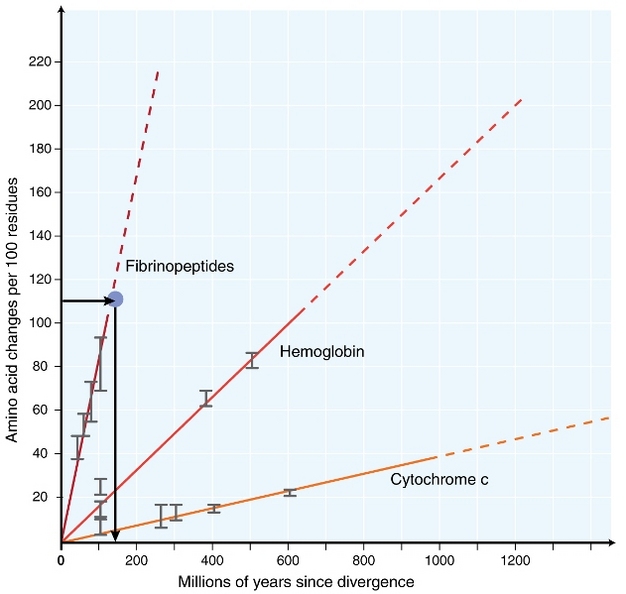
Suckerkandl& Pauling (1965) sammenlignede den konstante ophobning af aminosyresubstitutioner over tid med regelmæssige ‘flåter’ af ure og sagde, at ‘der kan eksistere et molekylært evolutionært ur’. Således blev udtrykket ‘molekylært ur’ oprindeligt opfundet for at beskrive ændringer i aminosyrer, der forekommer i proportional med tiden siden artsdivergens.
siden dets første anvendelse er udtrykket ‘molekylært ur’ blevet brugt i mange forskellige sammenhænge. I dag bruges det ofte til blot at henvise til antallet af ændringer eller ‘substitutioner’, akkumuleret i sekvensen af DNA eller proteiner i en given Slægt. Antallet af substitutioner pr. en defineret tidsenhed kan beskrives som’ hastigheden ‘for det molekylære ur, som i denne sammenhæng svarer til’evolutionær hastighed’. Bemærk, at den oprindelige konnotation stadig holder i nogle tilfælde. For eksempel, når folk siger ‘vi antog et molekylært ur’ (især i fylogenetiske analyser), betyder det, at antallet af substitutioner antages at forekomme konstant over tid.
begrebet et konstant molekylært ur har ekstraordinære konsekvenser for evolutionære biologer. Hvis et konstant molekylært ur som oprindeligt foreslået virkelig eksisterede, ville det at udlede timing af evolutionære begivenheder blive et ret ligetil problem (Figur 1). Det er imidlertid blevet helt klart, at substitutioner ikke forekommer konstant over tid i forskellige slægter (Kumar 2005). Ikke desto mindre har begrebet molekylært ur været ekstremt indflydelsesrig inden for molekylær evolution. En af de vigtigste ideer inspireret af begrebet molekylært ur er den neutrale teori om molekylær evolution.
det molekylære ur og neutralitet
da ideen om et konstant molekylært ur først opstod, blev det antaget, at den dominerende evolutionære kraft underliggende aminosyre-eller nukleotidsubstitutioner var naturlig selektion. Efter denne tankegang vil et konstant molekylært ur indikere, at adaptive substitutioner i forskellige arter forekommer konstant over tid. Det er imidlertid svært at forklare, hvordan adaptive substitutioner ville forekomme på en sådan urlignende måde. Teoretisk bestemmes skæbnen for adaptive mutationer af flere evolutionære parametre, såsom styrken af den selektive fordel ved denne mutation, størrelsen af den effektive population og adaptive mutationshastigheder (Kimura 1983). Disse parametre vil sandsynligvis variere mellem arter og endda inden for en art afhængigt af specifikke mutationer og deres interaktion med miljøer.
i stedet foreslog Kimura (1968, 1969), at de fleste ændringer på molekylært niveau har ringe funktionelle konsekvenser eller er ‘neutrale’. Hvis en mutation ikke har nogen form for konsekvens, bestemmes dens skæbne i befolkningen fuldstændigt ved tilfældig chance. Det betyder, at vi ikke kan forudsige, om en specifik neutral mutation i sidste ende vil blive rettet i befolkningen. Imidlertid kan den hastighed, hvormed neutrale substitutioner forekommer i befolkningen, forudsiges, fordi det afhænger af en enkelt parameter, nemlig mutationshastigheden (Kimura 1968).
lad os forestille os en befolkning med n antal haploide individer. Generation, vil det samlede antal mutationer i en generation være n gange u.da alle disse nye mutationer er neutrale, bestemmes deres skæbne fuldstændigt ved en tilfældighed. Med andre ord har alle mutationer lige chance for at nå fiksering (hvilket fører til en ‘substitution’). Sandsynligheden for, at hver ny neutral mutation vil nå fiksering, da der opstod en substitution, er simpelthen 1/N. Substitutionshastigheden beregnes som antallet af nye mutationer i hver generation (Nu) multipliceret med sandsynligheden for, at hver ny mutation når fiksering (1/N), hvilket er lig med u. med andre ord, for neutrale mutationer er substitutionshastigheden lig med mutationshastigheden!
derfor, hvis de fleste mutationer er neutrale (som foreslået i den neutrale teori), og hvis mutationshastigheder er konstante over tid, bør substitutioner også forekomme konstant over tid. Vi skal derefter observere urlignende regelmæssige substitutionshastigheder på molekylært niveau. Kimura (1969) betragtede således observationen af relativt konstant molekylært ur i proteinsekvenser som stærk støtte til den neutrale teori om molekylær evolution.
test af, hvordan det neutrale molekylære ur kører
ifølge den neutrale teori svarer spørgsmålet om, hvorvidt substitutionshastigheder er konstante over tid eller ej, til, om neutrale mutationshastigheder er konstante over tid. Af denne grund fokuserede mange efterfølgende undersøgelser på at analysere data fra neutrale steder for at afgøre, om neutrale mutationshastigheder faktisk er konstante over tid. Vi vil kort gennemgå, hvordan disse undersøgelser implementeres, inden vi diskuterer teoretiske debatter om molekylære ure. Analyser af proteinmolekylære ure fortsatte også, men debatterne omkring variation i proteinmolekylære ure er meget forskellige fra dem, der vedrører neutrale molekylære Ure, og vil ikke blive inkluderet i denne artikel. Interesserede læsere bør konsultere Gillespie (1991), Kumar (2005), Kim og Yi (2008) og Bedford et al. (2008).
de fleste empiriske analyser af neutrale molekylære ure er afhængige af sætningen om, at neutrale mutationshastigheder kan udledes af neutrale substitutionshastigheder (Kimura 1968, 1969). I praksis definerer hver undersøgelse en bestemt type steder i genomet som neutrale steder og sammenligner substitutionshastigheder for disse steder mellem slægter.
hvilke steder i genomet, der virkelig er neutrale, kan ikke bestemmes fuldstændigt, men forskere kom med flere nyttige fuldmagter. Før æraen med genomsekventering var de fleste tilgængelige sekvensdata dem fra proteinkodende DNA-sekvenser. Undersøgelser opdeler ofte proteinkodende DNA-sekvenser i to typer steder (Vu & Li 1985). Den første type steder inkluderer dem, for hvilke enhver ændring vil føre til aminosyresubstitutioner eller ‘ikke-synonyme steder’. Den anden type steder, ‘synonyme steder’, inkluderer dem, der koder for ‘degenererede’ positioner i kodontabellen, hvor en ændring ikke fører til en aminosyresubstitution. For eksempel koder TCT og TCC begge serin. Hvis den tredje position af disse kodoner ændres, vil de stadig producere de samme aminosyrer. Sådanne substitutioner ville være mindre synlige for naturlig udvælgelse. Derfor bør molekylære ure på synonyme steder være tættere på det neutrale molekylære ur end ikke-synonyme ure.
da sekventeringsteknikker avancerede, brugte nogle undersøgelser introner som fuldmagter til neutrale steder, da de ikke er inkorporeret i de modne mRNA ‘ er og dermed er mere tilbøjelige til at være neutrale (Yi et al. 2002). Sekvenser af inaktive transponerbare elementer, der blev indsat længe før artsdivergens, blev også ofte anvendt (disse kaldes ofte ‘forfædres gentagelser’, f.eks., 2003). Endelig anvendte nogle undersøgelser ikke-kodende DNA-sekvenser (alle sekvenser efter fjernelse af proteinkodende DNA-sekvenser) ekstraheret fra hele genomjusteringer for at teste neutrale molekylære ure (Elango et al. 2006, Huttley et al. 2007).
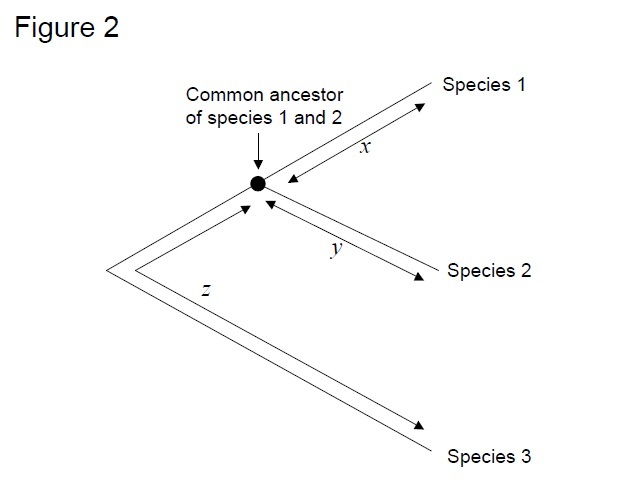
den mest anvendte test er den såkaldte ‘relative hastighedstest’ (Sarich & Vilson 1973). Tidsenhed estimeret ved at dividere det samlede antal forskelle (substitutioner) mellem proteiner af forskellige arter med divergenstiden, estimeret ud fra fossile optegnelser (Figur 1). Imidlertid, fossile poster er ikke tilgængelige for mange sammenligninger, og er forbundet med store fejlmarginer. Den relative hastighedstest overvinder nødvendigheden af fossile optegnelser (figur 2). Så længe der findes en udgruppesekvens til de to linjer af interesse, kan vi bestemme, om de to grene følger de samme eller forskellige molekylære ure uden kendskab til den absolutte tid for divergens (figur 2).
Hvad er determinanterne for neutrale molekylære ure?
næsten alle kontroverser i hjertet af debatter om neutrale molekylære ure stammer fra spørgsmålet om, hvad de vigtigste kilder til mutationer er. Dette spørgsmål er direkte relevant for forståelsen af mutationsmønstre, som er den ultimative kilde til evolutionær forandring og genetisk sygdom. Desuden er forståelse af, hvordan mutationshastigheder varierer mellem slægter og inden for genomer, et grundlæggende spørgsmål i komparativ genomik, der sigter mod at bruge sekvenssammenligninger til at identificere genomiske regioner, der er funktionelt vigtige.
så hvad bestemmer neutrale mutationshastigheder? En af de vigtigste bidragydere til neutrale molekylære ure er slægtsspecifik variation i generationstider. Fra tidligt blev ideen om et konstant neutralt molekylært ur opfattet som værende i strid med de molekylære mekanismer for kimlinjemutation. Det er længe blevet betragtet, at de fleste mutationer stammer fra fejl i DNA-replikation i kimlinjer (Haldane 1947, Muller 1954). Da mutationer opstår, når kimlinjens DNA replikeres til den næste generation, skal de akkumuleres i forhold til antallet af generationer snarere end den absolutte tid. Derfor, hvis vi sammenlignede antallet af substitutioner, der er akkumuleret i to slægter siden deres divergens, ville slægten med længere generationstid, efter at have gennemgået færre DNA-replikationshændelser, have færre substitutioner sammenlignet med slægten med den kortere generationstid. Derfor bør det molekylære ur køre langsommere i arter med længere generationstider. Denne ide kaldes ‘generation-time effect’.
faktisk blev genereringstidseffekten først observeret i studier af primater, selv før debatten om molekylært ur. Morris Goodman, der brugte immunologiske metoder til at undersøge artsrelateret på det tidspunkt, observerede, at den hastighed, hvormed nogle proteiner divergerer, syntes at være faldet i aber, især mennesker, sammenlignet med gamle verdens aber (Goodman 1961, 1962, 1963). Denne effekt kaldes ‘hominoid rate afmatning’. Da hominoider har længere generationstider end aber i den gamle verden, kan denne observation forklares med generationstidseffekten.
Vu& Li (1985) var de første til at teste hypotesen om genereringstidseffekt ved hjælp af DNA-sekvensdata. De brugte data fra 11 gener af primater og gnavere. Da primater har en meget længere generationstid end gnavere gør, bør molekyluret være hurtigere hos gnavere sammenlignet med primater. Faktisk fandt de, at gnavere for synonyme steder viser cirka to gange hastigheden af molekylær evolution sammenlignet med primater (Vu & Li 1985). For ikke-synonyme steder blev en sådan effekt imidlertid ikke fundet. Med andre ord, det neutrale molekylære ur, men ikke aminosyremolekylære ur, tikker hurtigere i gnaverlinjen sammenlignet med primatlinjen, hvilket passer godt til ideen om en generationstidseffekt.
efterfølgende undersøgelser gav yderligere støtte til hominoidhastigheden afmatning (Li& Tanimura 1987, Bailey et al. 1991) og hastighedsforskellen mellem gnaveren og primatlinjerne (Gu & Li 1992, Huttley et al. 2007). Desuden blev der observeret en hastighedsforskel i endnu mindre fylogenetiske skalaer, især hos primater: for eksempel kører det humane molekylære ur langsommere end chimpansens molekylære ur (Elango et al. 2006); satser i nye verdens aber er hurtigere end satserne i hominoider og gamle verdens aber (Steiper & Young 2006). De forskellige hastigheder af molekylære ure observeret i disse undersøgelser er kvalitativt i overensstemmelse med genereringstidseffekten.
de faktiske forskelle mellem slægter er imidlertid ikke kvantitativt i overensstemmelse med forskellen i generationstider. For eksempel viste Kumar & Subramanian (2002), at selvom forskellen i generationstider mellem primater og gnavere er meget større end forskellen mellem mennesker og aber i den gamle verden, er de observerede forskelle i molekylære ure ens i disse to sammenligninger. Det er værd at bemærke, at Kumar & Subramanian (2002) brugte specifikke statistiske filtre til at fjerne data, der viste ‘heterogene’ substitutionsmønstre, hvilket kunne have forårsaget en bias mod langsomt udviklende sekvenser (Yi et al. 2002). Ikke desto mindre forekommer forskellen i molekylære ure af primater og gnavere meget mindre end oprindeligt foreslået af VU & Li (1985). For eksempel Huttley et al. (2007) analyserede hele genomjusteringer af flere arter, herunder opossum, og viste, at hastighedsforskellen mellem eutherian-slægter og opossum-slægten (~30%) er meget større end hastighedsforskellen mellem menneskelige og mus-slægter (~14%). Disse eksempler viser, at graden af forskelle i molekylære ure varierer betydeligt mellem forskellige undersøgelser på grund af forskelle i datasæt og statistiske metoder. De viser også, at hastighedsforskelle mellem Slægter ikke helt kan redegøres for forskellen i generationstider alene. Det er klart, at der er andre bidragydere til neutrale molekylære ure.
faktisk synes andre livshistoriske træk end generationstider at variere med molekylære ure. Martin & Palumbi (1993) viste, at DNA-molekylære ure løber hurtigere i arter med lille kropsstørrelse. Denne observation førte til hypotesen om, at metaboliske hastigheder er vigtige determinanter for molekylære ure. En høj metabolisk hastighed producerer et stort antal mutagene iltradikaler, hvilket ville øge mutationshastighederne (Rand 1994). Fordi metaboliske hastigheder og kropsstørrelse generelt varierer med generationstider, har det været vanskeligt at skelne, hvilken af disse der udgør den vigtigste determinant for molekylære urhastigheder. Tsantes & Steiper (2009) har foreslået, baseret på data fra primater, at alder ved første reproduktion snarere end kropsstørrelse er den vigtigste determinant for molekylære ure. Da alder ved første reproduktion afspejler generationstidseffekten, understøtter denne undersøgelse ideen om, at generationstidseffekten er den vigtigste determinant for molekylært ur. Denne undersøgelse er dog stadig baseret på et begrænset antal slægter (fire par arter blev anvendt). Dermed, skelne mellem kropsstørrelse, genereringstidseffekt, og metaboliske hastigheder forbliver stadig som et vigtigt emne i generalisering og forståelse af neutrale molekylære ure.
desuden er vigtigheden af faktorer, der ikke varierer med generationstider, såsom DNA-methylering, blevet mere og mere værdsat. DNA-methylering er en kemisk modifikation af genomisk DNA, der findes i forskellige takser. I dyregenomer forekommer DNA-methylering næsten udelukkende ved cytosiner efterfulgt af guaniner (såkaldte ‘CpG’ er). Methylerede cytosiner har igen tendens til at mutere hurtigt til thyminer på grund af kemisk ustabilitet (Bird 1980). Faktisk forekommer mutationer forårsaget af DNA-methylering i det humane genom mere end en størrelsesorden oftere end andre mutationer (Nachman & krage 2000, Elango et al. 2008). Fordi mutationer forårsaget af DNA-methylering forekommer stort set uafhængigt af DNA-replikation, kan sådanne mutationer følge forskellige molekylære ure end andre. Specifikt kan mutationer forårsaget af DNA-methylering i stedet for genereringstidsafhængighed følge et tidsafhængigt molekylært ur, hvilket svarer til det, der oprindeligt blev foreslået af Cuckerkandl & Pauling (Kim et al. 2006)!
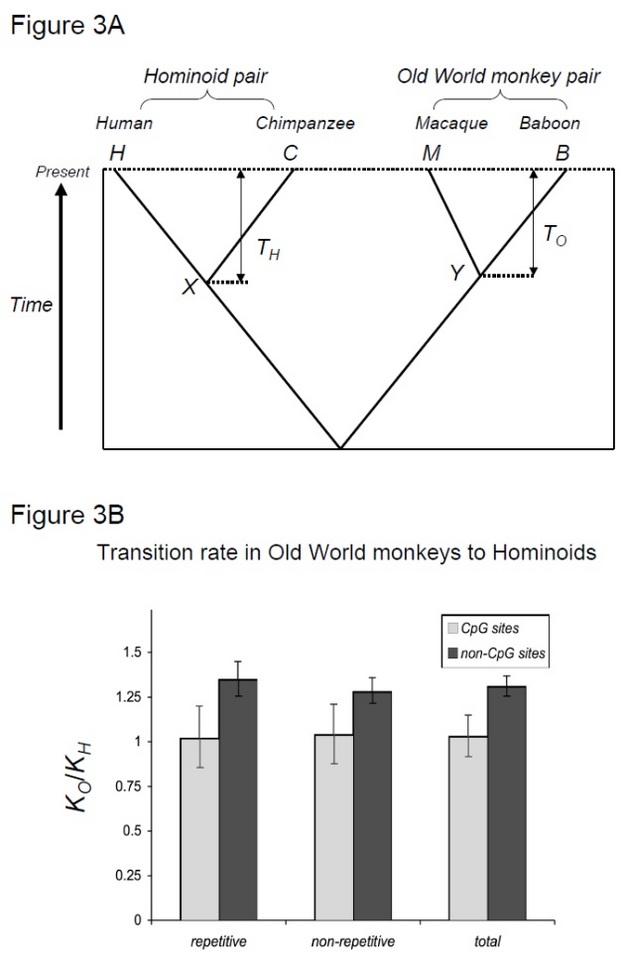
for at teste denne hypotese, Kim et al. (2006) sammenlignede human-chimpanse divergens med makak-bavian divergens, to arter par, der deler lignende divergenstider, men med forskellige generationstider (Steiper et al. 2004). Det Menneske-chimpanse-par (hominoid-paret) har meget længere generationstider sammenlignet med makak-bavian-paret (den gamle verdens abepar). Denne undersøgelse viste, at for ikke-CpG-steder akkumulerede det gamle verdens abepar cirka 30% flere substitutioner, hvilket kan forklares med den førnævnte hominoidhastighedsafmatningseffekt. I modsætning hertil viste molekylære ure på CpG-steder lignende antal substitutioner i hominoide og gamle verdens abepar (figur 3). Således eksisterer tidsafhængige og generationstidsafhængige molekylære ure inden for de samme genomer. Antagelsen om, at et enkelt molekylært ur kan eksistere for en given slægt, er ikke længere gyldigt, fordi de dominerende mutationskræfter varierer mellem genomiske regioner.
konklusioner
begrebet et konstant molekylært ur blev oprindeligt foreslået baseret på en begrænset mængde proteinsekvensdata. Selvom efterfølgende undersøgelser viste, at en sådan observation ikke er et generelt mønster i aminosyrer, har den haft betydelig indflydelse på området for molekylær evolution, især på udviklingen af den neutrale teori om molekylær evolution for DNA-sekvensdata. Efter den neutrale teori fokuserede undersøgelser på at belyse mønstre for variation i neutrale mutationshastigheder. I løbet af de sidste årtier, vi har observeret, at molekylære ure kører med forskellige hastigheder mellem slægter. Desuden kan variationsgraden variere afhængigt af de forskellige typer data og specifikke statistiske metoder, der anvendes. Genereringstidseffekten fortsætter med at holde på et kvalitativt niveau, men er utilstrækkelig til at forklare kvantitativ variation af neutrale mutationshastigheder blandt slægter. Livshistoriske træk og ikke-replikationsafhængige mutationer, såsom dem forårsaget af DNA-methylering, er også vigtige bidragydere til genomiske molekylære ure. Faktisk observeres forskellige typer molekylære ure selv inden for et genom, fordi de overvejende mutationsindgange varierer mellem forskellige genomiske regioner. Dermed, snarere end at antage et enkelt neutralt molekylært ur for hvert genom, fremtidige undersøgelser skal sigte mod at afsløre variationen af genomiske neutrale molekylære ure, at lære om genomiske mutationslandskaber. Sådanne oplysninger er ikke kun nyttige til forståelse af råmaterialet, der styrer molekylær udvikling og genetisk sygdom, men udgør også en kritisk komponent, der påvirker komparative og funktionelle genomiske analyser for at identificere funktionelle genomiske regioner.