Muskelplasticitet
skeletmuskulatur er det største væv i kroppen. Det er meget plastisk og kan gennemgå dramatiske ikke-patologiske svingninger i både masse og styrke som reaktion på et utal af miljøpåvirkninger. Efter modstandsøvelse eller eksponering for anabolske steroider gennemgår muskler hypertrofi og øger dens masse, tværsnitsareal og styrke (Aagaard, 2004; Schiaffino et al., 2013). Disse samme parametre kan falde kraftigt, når muskler indleder atrofi som reaktion på en hvilken som helst række fornærmelser, der inkluderer: sult, denervering, immobilisering, sepsis, kræftkakeksi, hjertesvigt, diabetes, nyresvigt, kronisk obstruktiv lungesygdom og erhvervet immundefektsyndrom (Schiaffino et al., 2013).
konsekvenserne af atrofi har indlysende konsekvenser for helbredet. Muskelsvaghed er en stor bidragyder til både dødelighed og sygelighed og er forbundet med den øgede risiko for alle dødsårsager (Metter et al., 2002). Faktisk kan reduktion af muskelatrofi i kræftkakeksi betydeligt forlænge livet (Jhou et al., 2010). Også mange ældre personer lider af sarkopeni, en langvarig muskelsvindforstyrrelse, der typisk begynder efter 50 år og indebærer et tab på omkring 1% af muskelmassen om året (bejle, 2017). Dette betyder, at sarkopeniske individer i en alder af 80 har mistet omkring 40% af deres muskelmasse, en nøglefaktor i fald, skrøbelighed og indlæggelser på plejehjem. Derfor er forståelsen af mekanismerne og potentielle terapeutiske reaktioner på atrofi af bred grundlæggende og klinisk interesse (Ali and Garcia, 2014; Cohen et al., 2015; Torben et al., 2017).
Myonuklear Domænehypotese
skeletmuskulatur er ret unik, fordi de modne celler er syncytiale og kan indeholde hundredvis af kerner. Dette er en nødvendig tilpasning, da individuelle muskelfibre kan være enorme og opnå længder på op til ~600 mm (23 tommer) (f.eks., 1998) og volumener, der kan være mere end 100.000 gange større end en typisk mononukleeret celle (Bruusgaard et al., 2003). Bidragene fra flere kerner er nødvendige for at producere store mængder mRNA, der er nødvendige for at styre den vidunderlige proteinsyntese, der kræves for at generere og vedligeholde det kontraktile apparat (Nevalainen et al., 2013).
både muskelens plastiske natur og dens syncytiale organisation har givet anledning til en kontrovers, der først for nylig ser ud til at være løst—den “myonukleære domænehypotese” (Kaisar og Larsson, 2014; Gundersen, 2016; Schvarts et al., 2016). Denne teori har sin oprindelse i begrebet “Virkungssph Kurrre” eller “indflydelsessfære” foreslået af Strassburger (1893), hvor han argumenterede for, at en kerne kun kan understøtte et diskret volumen af cytoplasma og således definere de øvre grænser for cellestørrelse. Dette princip blev uddybet yderligere af Gregory (2001), der hævdede, at cellulær deoksyribonukleinsyreindhold og volumen er tæt koblet. Skeletmuskulaturens syncytiale karakter hjælper disse celler med at overvinde denne begrænsning. Den myonukleære domænehypotese dikterer, at for at opretholde det korrekte nukleare til cytoplasmatiske forhold tilføjes nye kerner under hypertrofi og går tabt med atrofi.
der er betydelige data, der viser, at nukleart antal stiger med muskelhypertrofi (Moss, 1968; Cabric og James, 1983; Egner et al., 2016), selvom der stadig er en vis kontrovers (Murach et al., 2018). Denne proces har været et område med intens undersøgelse, og nogle af de underliggende molekylære mekanismer, der regulerer erhvervelsen af nye kerner under hypertrofi, er blevet defineret for nylig (Guerci et al., 2012; Torben et al., 2014; Ross et al., 2018). Disse supernumerære kerner erhverves, når stamcellebegrænsede stamceller, typisk satellitceller, smelter sammen med muskelfibre og bidrager med deres kerner (Brack and Rando, 2012; Bachman et al., 2018). Satellitceller befinder sig under kældermembranen og støder op til sarcolemma (1961; Mauro, 1961). De forbliver hvilende indtil stimuleret af enten anabolske steroider som testosteron eller ved fokal skade efter modstandstræning, på hvilket tidspunkt de genindtræder i cellecyklussen og spredes (Joubert og Tobin, 1995; Abreu et al., 2017). Nogle af dattercellerne smelter sammen med muskelfiberen og letter både reparation og vækst, mens andre arresterer og rekonstituerer satellitpuljen (Dumont et al., 2015; Goh og Millay, 2017). De resterende overskudsceller gennemgår apoptose (2008).
det kontroversielle aspekt af den myonukleære domænehypotese er påstanden om, at myonuklei går tabt under atrofi. Der er mange indgreb, der inducerer atrofi i dyremodeller, herunder: immobilisering, denervering og sepsis (Fitts et al., 1986; Minnaard et al., 2005; O ‘ Leary et al., 2012). I hvert tilfælde er der et nettotab af begge muskeltværsnitsareal (det primære assay) og udseendet af apoptotiske celler i vævet (McCall et al., 1998; Smith et al., 2000; Strasser et al., 2000; altid et al., 2003; McClung et al., 2007; Andrianjafiniony et al., 2010; Guo et al., 2012; Palumbo et al., 2012; Barnes et al., 2015; Cheema et al., 2015; Li et al., 2016; Torben et al., 2018). I disse undersøgelser måles apoptose i en række forskellige metoder, herunder caspase-aktivering, mitokondriel endogfrigivelse eller DNA-fragmentering . Disse undersøgelser giver overbevisende data om, at apoptose øges dramatisk i den tidlige fase af atrofi. For eksempel i en nylig omfattende undersøgelse (Guo et al., 2012), Guo et al. udsatte mus for 14 dages hindlimb-suspension, en behandling, der resulterede i en ~69% reduktion i muskelvådvægt og en ~43,8% reduktion i tværsnitsareal i forhold til den umanipulerede kontralaterale muskel. Forfatterne kvantificerede også et antal apoptosemarkører, herunder TUNELFARVNING, caspase-3 spaltning/aktivering og spaltning af Poly(adenosindiphosphat ribose) polymerase (PARP), et protein involveret i DNA-reparation. Hver af disse apoptoseforanstaltninger steg signifikant efter baglimb-suspension. Mens det primære fokus i dette papir var at evaluere den positive virkning af elektrisk stimulering på begrænsning af atrofi efter en fornærmelse, fortolker de og mange andre forskere denne slags data som en stærk støtte til den myonukleære domænehypotese.
fra et cellebiologisk perspektiv udgør det formodede tab af kerner i et syncytialvæv som skeletmuskulatur et stort praktisk problem. Hvordan kan en individuel kerne blive så kompromitteret, at dens genom hurtigt kondenserer og fragmenter, mens dens naboer vedvarer og hjælper med at opretholde muskelfiberens levedygtighed? I betragtning af at apoptose typisk medieres af aktiveringen af klassen af cysteinproteaser kendt som caspaser, er det ikke klart, hvilken mekanisme der kan tjene til at begrænse aktiviteten af en diffusibel protease inden for en fælles cytoplasma.
dette spørgsmål er blevet behandlet indirekte i en anden syncytialcelletype, den humane syncytiotrophoblast, et væv, der omgiver moderkagen og indeholder omkring 5-kilogram 1010-kerner (Mayhev et al., 1999). Når apoptose induceres i syncytiotrophoblasten, formerer den sig som en bølge med en hastighed på cirka 5 mikron pr., 2012). Derfor er der ingen” privilegerede ” regioner inden for syncytial cytoplasma, og alle kernerne ødelægges i sidste ende.
en af de vigtigste udfordringer med at analysere apoptose i skeletmuskulatur er, at det er et meget heterogent væv, hvor ca.halvdelen af dets kerner ligger uden for muskelfibre (Schmalbruch og Hellhammer, 1977). Disse mononukleerede celler inkluderer satellitceller, endotelceller, fibroblaster, pericytter og makrofager (Tedesco et al., 2010). Derfor er det meget vanskeligt at bestemme hvilken side af sarcolemma, en kerne er bosat, og dermed hvis det er en sand myonukleus.
In Vivo Time-Lapse Imaging af mærkede Musemuskelfibre
På trods af det store antal papirer, der demonstrerer apoptose under muskelatrofi, har flere forfattere stillet spørgsmålstegn ved disse resultater., 2002; Jong et al., 2005; Aravamudan et al., 2006; Gundersen og Bruusgaard, 2008; Duddy et al., 2011; Kaisar og Larsson, 2014). For eksempel ved anvendelse af isolerede muskelfibre in vitro, Duddy et al. bemærkede, at mens muskelfibervolumen faldt over tid, hvilket afspejler en atrofisk proces, syntes antallet af myonukleier at være stabilt (Duddy et al., 2011).
i en række elegante eksperimenter injicerede Gundersen lab individuelle ekstensorer digitorum longus (EDL) eller soleus muskler muskelfibre med farvestoffer, der uafhængigt mærkede kerner og cytoplasma i bedøvede mus (Bruusgaard og Gundersen, 2008; Bruusgaard et al., 2010). Dette gjorde det muligt for dem at identificere hver kerne inden for individuelle muskelfibre in vivo over tid og derefter evaluere dens skæbne. For eksempel blev EDL-muskler induceret til hypertrofi ved ablation af deres største synergister (Bruusgaard et al., 2010). Mellem dag 6 og 11 steg antallet af myonukleier med omkring 54%, og mellem dag 9 og 14 var der en 35% stigning i tværsnitsareal (Figur 1). Disse data er i overensstemmelse med hypotesen om, at muskler erhverver supernumerære kerner forud for den største vækst af fiberen under hypertrofi.

Figur 1. Myonuklei erhverves under hypertrofi, men går ikke tabt under atrofi hos mus. Mikrografer af samme EDL-muskelfiber over tid efter induktion af hypertrofi (øverste række) og den efterfølgende induktion af atrofi (nederste række). Fluorescerende mærkede oligonukleotider blev brugt til at visualisere kernerne in vivo. De stiplede linjer repræsenterer sarcolemma. Skala bar = 50 liter. (Tilpasset fra Bruusgaard et al., 2010. Brugt med tilladelse fra Proceedings of the National Academy of Sciences.)
de undersøgte derefter skæbnen for disse entydigt identificerbare myonukleier i de samme muskelfibre, da de blev induceret til at gennemgå atrofi. 50% reduktion i muskelfibervolumen, men intet tab af myonuklei (Figur 1; Bruusgaard et al., 2010). Manglende observation af nukleart tab skyldtes ikke den anvendte atrofiske stimulus, da de opnåede de samme resultater, da musklerne blev induceret til atrofi som reaktion på tetrodotoksininduceret nerveblokade, hindlimb suspension, kræft kakeksi eller detraining (Bruusgaard og Gundersen, 2008; Bruusgaard et al., 2010; Et Al., 2018a). Faktisk undersøgte de i løbet af deres studier mere end 200.000 individuelle myonukleier i atrofiske muskler og observerede kun 4 TUNEL-positive (apoptotiske) kerner, hvilket repræsenterer et tab på kun ~0,002% af kernerne (Bruusgaard et al., 2012). Disse data understøtter stærkt hypotesen om, at skeletmuskelatrofi ikke ledsages af myonuklear død.
Insektmuskel som Model
så kraftig som disse undersøgelser er, er der nogle begrænsninger forbundet med brugen af pattedyrsmodeller. For det første er de altid afhængige af eksperimentelle indgreb såsom denervering for at inducere muskelatrofi. For det andet bruges den kontralaterale muskel typisk som en intern kontrol, selvom den også udsættes for nogle af de samme miljøsignaler, såsom stresshormoner (Bonaldo og Sandri, 2013). Endelig kan skeletmuskler også gennemgå programmeret celledød (PCD) under udvikling, og disse modeller adresserer sjældent rollen som apoptose i disse celler.
en alternativ model, der ikke lider af nogen af disse begrænsninger, og tillader undersøgelse af muskelkerner under både atrofi og naturligt forekommende PCD, er den intersegmentale muskel (ISM) fra tobakshøgmoth Manduca seksta. ISMs er sammensat af plader af gigantiske muskelfibre, hvor hver celle er omkring 5 mm lang og op til 1 mm i diameter (figur 2a). ISMs fastgøres til segmentgrænserne i maven og genererer både larvernes krybende opførsel og eclosion (fremkomst) opførsel af den voksne møl, når den slipper ud af puppekutiklen i slutningen af metamorfose.
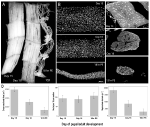
figur 2. Retention af myonuklei under både atrofi og død af de intersegmentale muskler (ISMs) fra møllen Manduca seksta. (A) ISMs fra tre udviklingsstadier: homeostatisk (dag 15 af pupal-voksen udvikling; venstre); atrofisk (dag 18; midten); og døende (18 timer efter eclosion; højre). Vægtstang er lig med ~1 mm (Tilpasset et al., 2016). (B) ISMs fra de samme udviklingsstadier blev ryddet og farvet med det nukleare farvestof DAPI og visualiseret via konfokal mikroskopi., 2016). (C) ISM-fiberafsnit (10 liter) blev farvet med det nukleare farvestof DAPI. Bemærk det dramatiske tab af muskelprotein (lysegrå område) under atrofi og død, men tilbageholdelsen af kerner i alle faser (tilpasset fra Schvarts et al., 2016). (D) kvantificering af ISM-fibervolumen (venstre), nukleart nummer (midten) og myonuklear domænestørrelse (højre) under homeostase, atrofi og død. (Betyder standard fejl.) (Tilpasset fra Schvarts et al., 2016).
på dag 15 af de normale 18 dage med pupal-voksenudvikling indleder ISMs et hormonelt udløst program for atrofi, der resulterer i et 40% tab af masse på tidspunktet for eclosion 3 dage senere (figur 2a; Truman og Truman, 1983). Dette dramatiske tab af muskelmasse svarer til det, der ses hos 80-årige personer med sarkopeni, men finder sted over 3 dage i stedet for 30 år. Mens ISMs mister masse, bevarer de normale fysiologiske egenskaber som hvilepotentiale og kraft/tværsnitsareal (2002). ISM ‘ erne bruges til at generere eclosion-adfærd i slutningen af dag 18 og derefter starte PCD, som begge udløses af et peptidhormon (1982, 1983). .
i modsætning til pattedyrsmuskler indeholder ISMs ikke kapillærer, satellitceller, endotelceller eller pericytter, hvilket betyder, at stort set alle kernerne i vævet befinder sig i selve muskelfibrene (Beaulaton og Lockshin, 1977). Vi brugte to uafhængige metoder til at overvåge nuklear skæbne under både atrofi og død i denne model., 2016). Den første var en standard anatomisk tilgang. Når ISMs gennemgår atrofi og død, er der dramatiske reduktioner i både volumen og tværsnitsareal af fibrene, men groft var nukleart antal uændret (figur 2b). Ved hjælp af sektionerede væv til kvantitative undersøgelser fandt vi,at der var en ~49% reduktion i fibertværsnitsareal under atrofi og et yderligere 30% fald i de tidlige stadier af døden (figur 2C, D; , 2016). Antallet af kerner ændrede sig imidlertid ikke i samme periode, hvilket betyder, at der funktionelt var en 84% reduktion i det myonukleære domæne.den anden metode, vi anvendte, var at måle DNA-indholdet af individuelle muskelfibre fra dyr på dag 13, når musklerne er homeostatiske, indtil 18 timer efter eclosion, når musklerne er stærkt degenererede., 2016). Vi kvantificerede DNA-indholdet af 420 individuelle celler og fandt ud af, at der ikke var noget signifikant tab under udviklingen. Disse biokemiske analyser stemmer Godt overens med de anatomiske data og antyder, at den relative koncentration af både kerner og genomisk DNA funktionelt steg, da musklerne atrofierede og døde. Følgelig antyder data fra både mus og møl, at muskelkerner ikke gennemgår apoptose under atrofi (eller PCD) og fører til den konklusion, at den myonukleære domænehypotese bør afvises.
konklusioner
disse observationer har en række implikationer for både forståelse af muskelens grundlæggende biologi og for udvikling af potentielle terapeutiske interventioner. Mens der er betydelige data, der rapporterer tilstedeværelsen af apoptotiske kerner i vævet efter atrofiske fornærmelser, antyder nylige data, at disse ikke er sande myonukleier, men snarere fordømte mononukleære celler, der ligger uden for muskelfiberen. Den primære årsag til denne uoverensstemmelse er, at de traditionelle værktøjer, der bruges til at detektere apoptose, mangler den opløsning, der kræves for tilstrækkeligt at bestemme, hvilken side af sarcolemma en døende celle befinder sig. Metoder, der specifikt mærker myonuklei, enten via in vivo injektioner af fluorescerende farvestoffer (Bruusgaard og Gundersen, 2008) eller via genetiske manipulationer til at udtrykke markørproteiner (Duddy et al., 2011), afslører få, hvis nogen apoptotiske kerner inden for de mærkede fibre. Den nylige demonstration af, at antistoffer rettet mod Protein Pericentriolært materiale 1 (PCM1) kun mærke autentisk myonuklei tilbyder et lovende værktøj, der kan anvendes på de traditionelle histologiske materialer, der rutinemæssigt analyseres i marken (Vinje et al., 2018b).
svigt af skeletmuskler til at anvende apoptose som en mekanisme til eliminering af myonuklei er ikke overraskende. Det ser ud til, at moden skeletmuskulatur stort set er udelukket fra at indlede apoptose, idet den opregulerer ekspressionen af potente overlevelsesproteiner som f.eks., 2009) og apoptose repressor med caspase recruitment domain (ARC)., 2011), samtidig med at man udtrykker lave niveauer af apoptosemediatorer som apoptotisk protease-aktiverende faktor 1 (Apaf-1) (Burgess et al., 1999). Dette giver mening teleologisk, da muskler er udsat for ekstreme forstyrrelser, herunder forstyrrelse af sarcolemma efter intens træning, så det ville være katastrofalt, hvis de udløste celledød snarere end initieret vævsreparation og muligvis hypertrofi. Ligesom andre terminalt differentierede celler har muskler en tendens til at anvende ikke-apoptotiske mekanismer, især autofagisk PCD (også kendt som type II degeneration) (Clarke, 1990., 1993; Kole et al., 2013; Ginet et al., 2014) når de dør under udvikling.
selvom der var en mekanisme, hvormed muskler selektivt kunne målrette individuelle kerner, er det ikke klart, at dette ville være gavnligt. Faktisk giver tilbageholdelsen af” overskydende ” kerner under atrofi en klar fordel for individet, da skeletmuskler ofte gennemgår cyklusser med atrofi og hypertrofi som reaktion på miljøforhold såsom tilgængelighed af fødevarer. Evnen til at komme sig hurtigt ved at udnytte allerede eksisterende myonukleier kan tjene en vigtig rolle i tilpasningen (Jackson et al., 2012) og hjælpe med at forklare fænomenet “muskelhukommelse” (Staron et al., 1991; Gundersen, 2016). Det er veldokumenteret inden for træningsfysiologi, at det er langt lettere at genvinde et vist niveau af muskelkondition gennem træning, end det var at opnå det først, selvom der har været en lang mellemliggende periode med detraining. Med andre ord er udtrykket” brug det eller tab det “måske mere præcist formuleret som” brug det eller tab det, indtil du arbejder på det igen.”Dette er blevet demonstreret direkte af et andet eksperiment fra Gundersen lab, der viste, at når en muskel har erhvervet nye kerner, bevarer den dem længe efter, at den hypertrofiske stimulus er fjernet. De inducerede muskelhypertrofi hos hunmus ved at behandle dem i 2 uger med testosteron og derefter undersøgt musklerne 3 uger efter steroid tilbagetrækning (Egner et al., 2013). Muskelvolumen var vendt tilbage til baseline, men de nyerhvervede kerner fortsatte endda 3 måneder senere. Når musklerne blev udsat for overbelastning for at genindføre hypertrofi, gennemgik de steroidbehandlede hurtigt en stigning på 36% i fibervolumen, mens kontrolmusklerne kun voksede med 6%. Disse data tyder på, at” overskydende ” kerner kunne mobiliseres hurtigt for at lette omskoling.
disse observationer har potentielle konsekvenser for folkesundhedspolitikken. Det har vist sig, at muskelvækst, fysiologisk funktion og regenerativ kapacitet alle falder med alderen, hovedsageligt på grund af reduceret satellitcelleproliferation (Blau et al., 2015). Derfor kan motion i ungdomsårene, når muskelvækst forbedres af hormoner, ernæring og en robust satellitpulje, funktionelt tjene til at give enkeltpersoner mulighed for at “bank” myonuklei, der kunne trækkes senere i livet for at bremse virkningerne af aldring og muligvis forhindre sarkopeni.
derudover har disse data konsekvenser inden for konkurrencesport. Brugen af anabolske steroider er en potent stimulans for muskel hypertrofi og tilsætning af nye myonuklei (Egner et al., 2013). Da disse kerner fortsætter længe efter steroid brug ender, atleter sandsynligvis udlede fordelene ved ulovlig stofbrug uden risiko for afsløring.
sammenfattende, mens tilføjelsen af nye kerner med muskelvækst stort set accepteres, kan det apoptotiske tab af kerner med atrofi ikke understøttes, hvilket antyder, at streng fortolkning af den myonukleære domænehypotese ikke kan understøttes. I stedet ser det ud til, at myonuklei, når den først er erhvervet, vedvarer, selv når en muskel bliver atrofisk eller indleder celledød.
Forfatterbidrag
forfatteren bekræfter at være den eneste bidragyder til dette værk og har godkendt det til offentliggørelse.
finansiering
arbejdet med denne publikation blev støttet af midler fra Eugene M. og Ronnie Isenberg professorat begavelse. Ingen corporate support blev ydet til forfatteren og ingen kommercielle produkter er nævnt eller godkendt.
interessekonflikt Erklæring
forfatteren erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
anerkendelser
forfatteren vil gerne takke Fru Christine Brun for hjælp til forberedelsen af dette manuskript og Eugene M. og Ronnie Isenberg professorat begavelse til støtte.
Aagaard, P. (2004). Gør musklerne “stærkere”: motion, ernæring, medicin. J. Musculoskelet. Neuronal. Interagere. 4, 165–174.
PubMed Abstract / Google Scholar
Abreu, P., Mendes, S. V., Ceccatto, V. M. Og Hirabara, S. M. (2017). Satellitcelleaktivering induceret af aerob muskeltilpasning som reaktion på udholdenhedstræning hos mennesker og gnavere. Livet Sci. 170, 33–40. doi: 10.1016 / j. lfs.2016.11.016
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Ali, S., og Garcia, J. M. (2014). Sarkopeni, kakeksi og aldring: diagnose, mekanismer og terapeutiske muligheder—en mini-gennemgang. Gerontologi 60, 294-305. doi: 10.1159 / 000356760
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
altid, S. E., Martyn, J. K., Ouyang, J., Chaudhrai, A. og Murlasits, S. S. (2003). Id2 expression during apoptosis and satellite cell activation in unloaded and loaded quail skeletal muscles. Am. J. Physiol. Regul. Integr. Comp. Physiol. 284, R540–R549. doi: 10.1152/ajpregu.00550.2002
PubMed Abstract | CrossRef Full Text | Google Scholar
Andrianjafiniony, T., Dupre-Aucouturier, S., Letexier, D., Couchoux, H., and Desplanches, D. (2010). Oxidative stress, apoptosis, and proteolysis in skeletal muscle repair after unloading. Am. J. Phys. Cell Physiol. 299, C307–C315. doi: 10.1152/ajpcell.00069.2010
PubMed Abstract / CrossRef fuldtekst / Google Scholar
Aravamudan, B., Mantilla, C. B., Han, V. S. og Sieck, G. C. (2006). Denerveringseffekter på myonuklear domænestørrelse af rottemembranfibre. J. Appl. Physiol. 100, 1617–1622. doi: 10.1152 / japplphysiol.01277.2005
CrossRef fuldtekst/Google Scholar
Bachman, J. F., Klose, A., Liu, V., Paris, N. D., Blanc, R. S., M., et al. (2018). Præpubertal skeletmuskelvækst kræver 7-udtrykker satellitcelleafledt myonukleært bidrag. Udvikling 145: 167197.
Google Scholar
Barnes, B. T., Confides, A. L., Rich, M. M. og Dupont-Versteegden, E. E. (2015). Forskellige muskelapoptotiske veje aktiveres i muskler med forskellige fibertyper i en rottemodel af kritisk sygdom myopati. J. Muskel Res. Celle Motil. 36, 243–253. doi: 10.1007 / s10974-015-9410-8
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Beaulaton, J., og Lockshin, R. A. (1977). Ultrastrukturel undersøgelse af den normale degeneration af de intersegmentale muskler i Anthereae polyphemus og Manduca seksta (insecta, lepidoptera) med særlig henvisning til cellulær autofagi. J. Morphol. 154, 39–57. doi: 10.1002 / jmor.1051540104
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Bentsinger, C. F., von Maltsahn, J., Dumont, N. A., Stark, D. A., Vang, Y. K., Nhan, K., Et Al. (2014). Vnt 7A stimulerer MyoGen stamcellemotilitet og engraftment, hvilket resulterer i forbedret muskelstyrke. J. Cell Biol. 205, 97–111. doi: 10.1083 / jcb.201310035
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Blau, H. M., Cosgrove, B. D. og Ho, A. T. (2015). Den centrale rolle muskelstamceller i regenerativ svigt med aldring. Nat. Middelhavs. 21, 854–862. doi: 10.1038 / nm.3918
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Bonaldo, P., og Sandri, M. (2013). Cellulære og molekylære mekanismer for muskelatrofi. Dis. Model. Mech. 6, 25–39. doi: 10.1242 / dmm.010389
CrossRef fuldtekst / Google Scholar
Brack, A. S. og Rando, T. A. (2012). Vævsspecifikke stamceller: lektioner fra skeletmuskel satellitcelle. Cellestamcelle 10, 504-514. doi: 10.1016 / j. stem.2012.04.001
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Bruusgaard, J. C., Egner, I. M., Larsen, T. K., Dupre-Aucouturier, S., Desplanches, D. og Gundersen, K. (2012). Ingen ændring i myonukleært nummer under muskelaflæsning og genindlæsning. J. Appl. Physiol. 113, 290–296. doi: 10.1152 / japplphysiol.00436.2012
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Bruusgaard, J. C., Og Gundersen, K. (2008). In vivo time-lapse mikroskopi afslører intet tab af Murin myonuklei i uger med muskelatrofi. J. Clin. Investere. 118, 1450–1457. doi: 10.1172 / JCI34022
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Bruusgaard, J. C., Johansen, I. B., Egner, I. M., Rana, A. A. og Gundersen, K. (2010). Myonuklei erhvervet ved overbelastningsøvelse forud for hypertrofi og går ikke tabt ved detraining. Proc. Natl. Acad. Sci. U. S. A. 107, 15111-15116. doi: 10.1073 / pnas.0913935107
CrossRef fuldtekst / Google Scholar
Bruusgaard, J. C., Liestol, K., Ekmark, M., Kollstad, K. og Gundersen, K. (2003). Antal og rumlig fordeling af kerner i muskelfibrene hos normale mus undersøgt in vivo. J. Physiol. 551, 467–478. doi: 10.1113 / jphysiol.2003.045328
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Burgess, D. H., Svensson, M., Dandrea, T., Gronlund, K., Hammarkvist, F., Orrenius, S., et al. (1999). Humane skeletmuskelcytosoler er ildfaste over for cytokrom C-afhængig aktivering af type II caspaser og mangler APAF-1. Celledød Er Forskellig. 6, 256–261. doi: 10.1038 / sj.cdd.4400489
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Cabric, M., and James, N. T. (1983). Morfometriske analyser på musklerne i trænede og utrænede hunde. Er. J. Anat. 166, 359–368. doi: 10.1002 / aja.1001660309
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Cheema, N., Herbst, A., Mckenie, D. og Aiken, J. M. (2015). Apoptose og nekrose medierer tab af skeletmuskulatur i aldersinducerede mitokondrielle abnormiteter. Aldrende Celle 14, 1085-1093. doi: 10.1111 / acel.12399
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Clarke, P. G. (1990). Udviklingscelledød: morfologisk mangfoldighed og flere mekanismer. Anat. Embryol. 181, 195–213.
Google Scholar
Cohen, S., Nathan, J. A. og Goldberg, A. L. (2015). Muskelsvind i sygdom: molekylære mekanismer og lovende terapier. Nat. Rev. Drug Discov. 14, 58–74. doi: 10.1038 / nrd4467
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Duddy, J. J., Cohen, T., Dugues, S. og Partridge, T. A. (2011). Den isolerede muskelfiber som model for ubrugt atrofi: karakterisering ved hjælp af PhAct, en metode til at kvantificere f-actin. Eksp. Celle Res. 317, 1979-1993. doi: 10.1016 / j.2011.05.013
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Dumont, N. A., Vang, Y. H. og Rudnicki, M. A. (2015). Interne og eksterne mekanismer, der regulerer satellitcellefunktion. Udvikling 142, 1572-1581. doi: 10.1242 / dev.114223
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Egner, I. M., Bruusgaard, J. C., Eftestol, E. og Gundersen, K. (2013). En cellulær hukommelsesmekanisme hjælper overbelastningshypertrofi i muskler længe efter en episodisk eksponering for anabolske steroider. J. Physiol. 591, 6221–6230. doi: 10.1113 / jphysiol.2013.264457
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Egner, I. M., Bruusgaard, J. C. og Gundersen, K. (2016). Satellitcelleudtømning forhindrer fiberhypertrofi i skeletmuskulatur. Udvikling 143, 2898-2906. doi: 10.1242 / dev.134411
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Fitts, R. H., Metger, J. M., Riley, D. A., B. R. (1986). Modeller af ubrugt: en sammenligning af baglimb suspension og immobilisering. J. Appl. Physiol. 60, 1946–1953.
Google Scholar
Ginet, V., Pittet, M. P., Rummel, C., Osterheld, M. C., Meuli, R., Clarke, P. G., et al. (2014). Døende neuroner i thalamus af kvælte sigt nyfødte og rotter er autofagiske. Ann. Neurol. 76, 695–711. doi: 10.1002 / ana.24257
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Goh, K., og Millay, D. P. (2017). Krav til myomaker-medieret stamcellefusion til skeletmuskelhypertrofi. elife 6. doi: 10.7554 / eLife.20007
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Gregory, T. R. (2001). Tilfældighed, coevolution eller årsagssammenhæng? DNA-indhold, cellestørrelse og C-værdien enigma. Biol. Pastor Camb. Philos. Soc. 76, 65–101. doi: 10.1017/s1464793100005595
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Guerci, A., Lahoute, C., Hebrard, S., Collard, L., Graindorge, D., Favier, M., et al. (2012). Srf-afhængige paracrine signaler produceret af myofibre styrer satellitcellemedieret skeletmuskelhypertrofi. Cellemetab. 15, 25–37. doi: 10.1016 / j. cmet.2011.12.001
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Gundersen, K. (2016). Muskelhukommelse og en ny cellulær model for muskelatrofi og hypertrofi. J. Eksp. Biol. 219, 235–242. doi: 10.1242 / jeb.124495
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Gundersen, K., og Bruusgaard, J. C. (2008). Nukleare domæner under muskelatrofi: kerner tabt eller paradigme tabt? J. Physiol. 586, 2675–2681. doi: 10.1113 / jphysiol.2008.154369
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Guo, B. S., Cheung, K. K., Yeung, S. S., Jang, B. T. og Yeung, E. V. (2012). Elektrisk stimulering påvirker satellitcelleproliferation og apoptose ved losning-induceret muskelatrofi hos mus. PLoS en 7: e30348. doi: 10.1371 / tidsskrift.pone.0053523
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Jackson, J. R., Mula, J., Kirby, T. J., Fry, C. S., Lee, J. D., Ubele, M. F., et al. (2012). Satellitcelleudtømning hæmmer ikke voksen skeletmuskulaturgenvækst efter losseinduceret atrofi. Er. J. Phys. Cellefysiol. 303, C854-C861. doi: 10.1152 / ajpcell.00207.2012
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Joubert, Y., og Tobin, C. (1995). Testosteronbehandling resulterer i, at hvilende satellitceller aktiveres og rekrutteres til cellecyklus i rotte levator ani muskel. Dev. Biol. 169, 286–294. doi: 10.1006 / dbio.1995.1144
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
kat, B. (1961). Afslutningen af den afferente nervefiber i frøens muskelspindel. Philos. Trans. R. Soc. Lond. B Biol. Sci. 243, 221–240.
PubMed abstrakt/Google Scholar
Kletsien, H., Hare, A. J., Leverson, G. og Connor, N. P. (2018). Aldersrelateret effekt af celledød på fibermorfologi og antal i tunge muskler. Muskel Nerve 57, E29-E37. doi: 10.1002 / mus.25671
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Kole, A. J., Annis, R. P. og Deshmukh, M. (2013). Modne neuroner: udstyret til overlevelse. Celledød Dis. 4: e689. doi: 10.1038 / cddis.2013.220
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Li, T. S., Shi, H., Vang, L. og Yan, C. (2016). Effekt af knoglemarv mesenkymale stamceller på satellitcelleproliferation og apoptose ved immobiliseringsinduceret muskelatrofi hos rotter. Middelhavs. Sci. Monit. 22, 4651–4660. doi: 10.12659 / MSM.898137
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Lockshin, R. A., C. M. (1965). Programmeret celledød-I. cytologi af degeneration i de intersegmentale muskler i pernyi silkmoth. J. Insekt Physiol. 11, 123–133. doi: 10.1016/0022-1910(65)90099-5
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Longtine, M. S., Barton, A., Chen, B. og Nelson, D. M. (2012). Levende celleafbildning viser, at apoptose initierer lokalt og formerer sig som en bølge gennem syncytiotrophoblaster i primære kulturer af humane placentale villøse trofoblaster. Placenta 33, 971-976. doi: 10.1016 / j. placenta.2012.09.013
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Mauro, A. (1961). Satellitcelle af skeletmuskelfibre. J. Biophys. Biochem. Cytol. 9, 493–495.
Google Scholar
Mayhe, T. M., Leach, L., McGee, R., Ismail, Myklebust, R. og Lammiman, M. J. (1999). Proliferation, differentiering og apoptose i villøs trofoblast ved 13-41 ugers drægtighed (inklusive observationer af annulerede lameller og nukleare porekomplekser). Placenta 20, 407-422. doi: 10.1053 / sted.1999.0399.
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
McCall, G. E., Allen, D. L., Linderman, J. K., Grindeland, R. E., Roy, R. R., Mukku, V. R., et al. (1998). Vedligeholdelse af myonuklear domænestørrelse i rotte soleus efter overbelastning og væksthormon/IGF-i behandling. J. Appl. Physiol. 84, 1407–1412.
Google Scholar
McClung, J. M., Deruisseau, K. C., Falk, D. J., Deering, M. A., Lee, Y., et al. (2007). Caspase – 3 regulering af membran myonukleært domæne under mekanisk ventilationsinduceret atrofi. Er. J. Respir. Crit. Pleje Med. 175, 150–159. doi: 10.1164 / rccm.200601 – 142oc
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Metter, E. J., Talbot, L. A., Schrager, M.( 2002). Skeletmuskelstyrke som en forudsigelse for dødelighed af alle årsager hos raske mænd. J. Gerontol. En Biol. Sci. Middelhavs. Sci. 57, B359-B365. doi: 10.1093 / gerona / 57.10.B359
PubMed Abstract / CrossRef fuldtekst / Google Scholar
Minnaard, R., Drost, M. R., A. J., van Kranenburg, G. P., Kuipers, H. og Hesselink, M. K. (2005). Skeletmuskulatur spilde og kontraktile ydeevne i septisk rotter. Muskel Nerve 31, 339-348. doi: 10.1002 / mus.20268
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Moss, F. P. (1968). Forholdet mellem fibrenes dimensioner og antallet af kerner under normal vækst af skeletmuskulatur hos tamhøns. Er. J. Anat. 122, 555–563. doi: 10.1002 / aja.1001220308
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Murach, K. A., Englund, D. A., Dupont-Versteegden, E. E., McCarthy, J. J. og Peterson, C. A. (2018). Myonuklear domænefleksibilitet udfordrer stive antagelser om satellitcellebidrag til skeletmuskelfiberhypertrofi. Front. Physiol. 9:635. doi: 10.3389 / fphys.2018.00635
CrossRef fuldtekst / Google Scholar
Nevalainen, M., Kaakinen, M. Og Metsikko, K. (2013). Fordeling af mRNA-udskrifter og oversættelsesaktivitet i skeletmyofibre. Cellevæv Res. 353, 539-548. doi: 10.1007 / s00441-013-1659-Google Scholar
PubMed Abstract | CrossRef fuldtekst/Google Scholar
O ‘ Leary, M. F., Vainshtein, A., Carter, H. N., Jang, Y. og Hood, D. A. (2012). Denerveringsinduceret mitokondriel dysfunktion og autofagi i skeletmuskulatur hos apoptose-mangelfulde dyr. Er. J. Phys. Cellefysiol. 303, C447-C454. doi: 10.1152/ajpcell.00451.2011
PubMed Abstract | CrossRef Full Text | Google Scholar
Palumbo, C., Rovesta, C., and Ferretti, M. (2012). Striated muscle fiber apoptosis after experimental tendon lesion in a rat model. J. Anat. 221, 358–363. doi: 10.1111/j.1469-7580.2012.01554.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Qaisar, R., and Larsson, L. (2014). What determines myonuclear domain size? Indian J. Physiol. Pharmacol. 58, 1–12.
PubMed Abstract | Google Scholar
Ross, J. A., Levy, Y., Svensson, K., Philp, A., Schenk, S. og Ochala, J. (2018). SIRT1 regulerer atomnummer og domænestørrelse i skeletmuskelfibre. J. Cell. Physiol. 233, 7157–7163. doi: 10.1002 / jcp.26542
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Schiaffino, S., Dyar, K. A., Ciciliot, S., Blaau, B. Og Sandri, M. (2013). Mekanismer, der regulerer skeletmuskulatur vækst og atrofi. FEBS J. 280, 4294-4314. doi: 10.1111 / febs.12253
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Schmalbruch, H. og Hellhammer, U. (1977). Antallet af kerner i voksne rotte muskler med særlig henvisning til satellitceller. Anat. Rec. 189, 169–175. doi: 10.1002 / ar.1091890204
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
L. M. (2008). Atrofi og programmeret celledød af skeletmuskulatur. Celledød Er Forskellig. 15, 1163–1169. doi: 10.1038 / cdd.2008.68
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
brun, C., McLaughlin, K., Smith, V. og Bigelav, C. (2016). Det myonukleære domæne opretholdes ikke i skeletmuskulatur under hverken atrofi eller programmeret celledød. Er. J. Phys. Cellefysiol. 311, C607-C615. doi: 10.1152 / ajpcell.00176.2016
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Schvarts, L. M. og Ruff, R. L. (2002). Ændringer i kontraktile egenskaber af skeletmuskulatur under udviklingsmæssigt programmeret atrofi og død. Er. J. Phys. Cellefysiol. 282, C1270-C1277. doi: 10.1152 / ajpcell.01275.2000
CrossRef Fuld tekst/Google Scholar
L. M., Smith, S. V., Jones, M. E. Osborne, B. A. (1993). Forekommer alle programmerede celledødsfald via apoptose? Proc. Natl. Acad. Sci. USA 90, 980-984.
Google Scholar
L. M. Og Truman, J. V. (1983). Hormonal kontrol af metamorfe udviklingshastigheder i tobakshornorm Manduca seksta. Dev. Biol. 99, 103–114. doi: 10.1016/0012-1606(83)90257-9
PubMed Abstract | CrossRef fuldtekst | Google Scholar
PubMed Abstract / CrossRef fuldtekst / Google Scholar (1982). Peptid og steroid regulering af muskel degeneration i et insekt. Videnskab 215, 1420-1421. doi: 10.1126 / videnskab.6278594
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Smith, H. K., L., Martyn, J. A. og Bass, J. J. (2000). Nuklear DNA-fragmentering og morfologiske ændringer i voksen kanin skeletmuskulatur efter kortvarig immobilisering. Cellevæv Res. 302, 235-241. doi: 10.1007 / s004410000280
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Smith, M. I., Huang, Y. Y. og Deshmukh, M. (2009). Skeletmuskeldifferentiering fremkalder endogen HSIAP for at begrænse den apoptotiske vej. PLoS en 4: e5097. doi: 10.1371 / tidsskrift.pone.0005097
CrossRef Fuld tekst/Google Scholar
Staron, R. S., Leonardi, M. J., Karapondo, D. L., Malicky, E. S., Falkel, J. E., Hagerman, F. C., Et Al. (1991). Styrke-og skeletmuskulaturtilpasninger hos tunge modstandstrænede kvinder efter detraining og omskoling. J. Appl. Physiol. 70, 631-640.
Google Scholar
Strassburger, E. (1893). Kernens virkefelt og cellestørrelsen. Histol Beitr. 5, 97-124.
Google Scholar
Strasser, H., Tiefenthaler, M., Steinlechner, M., Eder, I., Bartsch, G. og Konvalinka, G. (2000). Alder afhængig apoptose og tab af rhabdosphincter celler. J. Urol. 164, 1781–1785. doi: 10.1016 / S0022-5347 (05)67106-6
PubMed Abstract | CrossRef fuldtekst | Google Scholar
Tedesco, F. S., Dellavalle, A., Dia-manera, J., Messina, G. og Cossu, G. (2010). Reparation af skeletmuskulatur: regenerativt potentiale i skeletmuskelstamceller. J. Clin. Investere. 120, 11–19. doi: 10.1172 / JCI40373
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Vada, K. I., Takahashi, H., Katsuta, S. og soja, H. (2002). Intet fald i myonukleært antal efter langvarig denervering hos modne mus. Er. J. Phys. Cellefysiol. 283, C484-C488. doi: 10.1152 / ajpcell.00025.2002
CrossRef fuldtekst / Google Scholar
Vinje, I. M., Sheng, Hansson, K. A., Solbrr, A., Tennrite, S., Saatcioglu, F., et al. (2018a). Kakeksi inducerer ikke tab af myonuklei eller muskelfibre under røntgenfotograferet prostatacancer hos mus. Acta Physiol. e13204. doi: 10.1111 / apha.13204
CrossRef fuldtekst/Google Scholar
Vinje, I. M., Bengtsen, M., Eftestol, E., Juvkam, I., Bruusgaard, J. C. og Gundersen, K. (2018b). Specifik mærkning af myonuklei af et antistof mod pericentriolært materiale 1 på skeletmuskelvævssektioner. Acta Physiol. 223: e13034. doi: 10.1111 / apha.13034
CrossRef Fuld tekst / Google Scholar
bejle, J. (2017). Sarkopeni. Clin. Geriatr. Middelhavs. 33, 305–314. doi: 10.1016 / j. cger.2010.02.012
CrossRef Fuld tekst/Google Scholar
JJ, R., Ferry, A. L., og Dupont-Versteegden, E. E. (2011). Celledød-resistens af differentierede myotubes er forbundet med forbedrede anti-apoptotiske mekanismer sammenlignet med myoblaster. Apoptose 16, 221-234. doi: 10.1007 / s10495-010-0566-9
PubMed abstrakt / CrossRef Fuld tekst/Google Scholar
Yang, D., Morris, S. F. og Sigurdson, L. (1998). Sartorius-muskelen: anatomiske overvejelser for rekonstruktive kirurger. Surg. Radiol. Anat. 20, 307–310. doi: 10.1007
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Jong, H., Roy, R. R., Siengthai, B. og Edgerton, V. R. (2005). Virkninger af inaktivitet på fiberstørrelse og myonukleært antal i rotte soleus muskel. J. Appl. Physiol. 99, 1494–1499. doi: 10.1152 / japplphysiol.00394.2005
CrossRef Fuld tekst/Google Scholar
J. L., Lu, J., sang, Y., K. S., Jiao, et al. (2010). Reversering af kræftkakeksi og muskelsvind ved Act RIIB-antagonisme fører til langvarig overlevelse. Celle 142, 531-543. doi: 10.1016 / j.celle.2010.07.011
PubMed Abstract / CrossRef fuldtekst / Google Scholar
Silaaldini, M. M., Marsetti, E., Picca, A. og Murlasits, å. (2017). Biokemiske veje for sarkopeni og deres modulering ved fysisk træning: en fortællende gennemgang. Front. Middelhavs. 4:167. doi: 10.3389 / fmed.2017.00167
CrossRef Fuld tekst / Google Scholar