introduktion
International conservation policy anerkender biodiversitet på tre niveauer, økosystem, arter og genetiske, og at forvaltningen bør sigte mod at bevare alle tre (konvention om biologisk mangfoldighed, 2007). Dette afspejles tydeligt i konventionen om biologisk mangfoldighed, Aichi biodiversitetsmål, der blev vedtaget i 2010 (https://www.cbd.int/sp/targets/), hvor der er specifik reference i mål og mål, ikke kun til økosystemer og arter, men også til genetisk mangfoldighed. Alligevel er de nuværende tilgange til bevarelse af biodiversitet stort set baseret på geografiske områder, økosystemer, økologiske samfund og arter med mindre opmærksomhed på genetisk mangfoldighed og artspopulationen kontinuum. Faktisk fokuserer flertallet af bevaringsudøvere på arter som måling af biodiversitet, med en vis overvejelse af intraspecifikke enheder inden for intensivt forvaltede truede arter. I denne sammenhæng er der et naturligt ønske om at anvende enkle kategoriseringer—navngivne arter og andre navngivne takser (dvs.underarter) og undertiden bevaringsenheder inden for arter. Arter giver især et fælles mål for biodiversitet, der understøtter meget videnskabelig og ledelsesmæssig indsats (Mace, 2004, 2017). Som en grundlæggende bevaringsenhed bruges arter ofte til at kvantificere biodiversitetsværdi gennem lister over arter inden for beskyttede områder, identifikation af truede arter inden for jurisdiktioner og som grundlag for biosikkerhedsprocedurer. Derudover er arter typisk de enheder, som vi kommunikerer bevaringsspørgsmål med til offentligheden.
denne impuls til blot at identificere diskrete enheder i naturen strider mod flere relaterede virkeligheder. For det første er speciering ikke øjeblikkelig. Det er snarere typisk en langvarig proces (Dynesius and Jansson, 2014; et al., 2016), der udvikler sig fra isolerede populationer, måske underlagt forskellige selektionstryk, til entydige arter over lange tidsskalaer med hyppige tilbageførsler forårsaget af miljøændringer eller simpel genetisk sammensmeltning, når intervaller forbindes igen (Figur 1; Seehausen et al., 2008; Rosenblum et al., 2012). For det andet er de fleste anerkendte arter ikke genetisk ensartede. Divergens (drift og langvarig isolation) og retikulation (genstrømning, hybridisering, introgression og rekombination) former effektivt geografisk og genomisk variation inden for arter (Cutter and Payseur, 2013; , 2016), og nogle arter kan struktureres i lange isolerede populationer, hvoraf nogle kan berettige anerkendelse som fulde, men måske morfologisk kryptiske arter (Bickford et al., 2007; Jorger og Schr Kristdl, 2013). For det tredje, så længe anerkendt, er navngivne arter ikke ækvivalente med hensyn til hvor meget evolutionær arv de repræsenterer—enkelt sagt kan en platypus (som repræsentant for en gammel divergeret Slægt) betragtes som mere værd end en for nylig divergeret gnaver (Isaac et al., 2007). For det fjerde er artsbetegnelser ikke støbt i sten, snarere er de hypoteser, der skal testes, når nye beviser bliver tilgængelige (Hey et al., 2003). Hvert af disse spørgsmål skaber kompleksitet og tvetydighed i afgrænsning af arter generelt og tildeling af bevaringsopmærksomhed til dem eller deres komponentpopulationer. Derudover er afgrænsningen af arter også underlagt forskellige artskoncepter og måder at diagnosticere dem på, hvilket har forårsaget stor foruroligelse blandt bevaringsbiologer (Mace, 2004; Frankham et al., 2012; Garnett og Christidis, 2017).
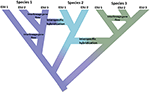
Figur 1. Speciering som et kontinuum, der udvikler sig fra isolerede populationer til entydige arter vist ved skiftet i farve med farverne, der repræsenterer forskellige arter. Nogle slægter, der opstår på grund af befolkningsisolering, udvikler sig til ægte arter med træets indre knudepunkter, der repræsenterer initiering af specieringsprocessen. Andre slægter udvikler sig ikke til ægte arter, men afviger over tid, hvilket indikerer, at arter ikke er genetisk ensartede. Disse slægter er vist som evolutionært signifikante enheder (ESU ‘ er). Genstrømning mellem afstamning er indiceret for Art 1 og Art 3. I dette tilfælde kan evolutionær uafhængighed være forbigående, fordi to slægter, der tilhører den samme art, ikke er reproduktivt isoleret. Mens de forbliver som forskellige slægter (arter), kan interspecifik hybridisering forekomme med begrænset genudveksling (skift i farve) mellem arter.implementering af effektive bevaringsstrategier, der sigter mod at beskytte både arter og genetisk mangfoldighed, vil drage fordel af både et mere konsistent syn på, hvordan vi afgrænser arter og andre enheder på tværs af bestandsartskontinuumet, og hvordan disse enheder fortolkes i forskellig lovgivning globalt. Nye kapaciteter fra genomics giver både muligheder og udfordringer for at nå disse mål. I det følgende overvejer vi: (i) hvordan genomskalaanalyser kan forenes med en konservativ tilgang til afgrænsning og navngivning af arter, samtidig med at de identificerer hovedkomponenter i genetisk mangfoldighed inden for arter, (ii) hvordan eksisterende bevaringslovgivning på tværs af flere kontinenter, der definerer “arter” til at omfatte en bred vifte af infrarøde kategorier, kan bruges i betragtning af den høje opløsning, der tilbydes af genomskærme, og (iii) hvordan indsigten i specieringsprocesser og strukturering af variation fra genomik stemmer overens med lovgivningen og derefter til styring. Vi foreslår derefter nogle måder fremad i at tilpasse bevaringsenheder med lovgivningsmæssige tilgange for at beskytte det fulde spektrum af mangfoldighed fra populationer gennem arter.
genomik, speciering og taksonomi
ligesom mange områder inden for biologisk videnskab, systematik, praksis med at navngive taksa og forstå deres forhold og hvordan de dannes, bliver betydeligt avanceret af genomik og statistiske modeller. Uanset om det er gennem helgenomsekventering (f.eks., 2017) eller genomprøvetagning (SNP ‘ er, target capture-sekventering; Jones and Good, 2016; Leache and Oaks, 2017), har vi nu en hidtil uset opløsning af mønstre af genetisk mangfoldighed (Funk et al., 2012). Sammen med stadig stærkere statistiske modeller til at udlede historier om genomisk og artsdivergens, disse data giver ny indsigt i de evolutionære processer, der genererer arter og genetisk variation.
anvendelse af disse nye værktøjer genererer to modsatte indsigter. På den ene side afslører mange undersøgelser meget divergerende genetiske populationer inden for navngivne arter, såkaldte kryptiske arter (Bickford et al., 2007; Jorger og Schr Kristdl, 2013; Struck et al., 2017). Dette er ikke noget nyt, da det bygger på en lang historie med rumlig befolkningsgenetik og molekylær fylogeografi, der har været en produktiv indsats i tre årtier (avis et al., 1987). Hvad genomik bringer til denne bestræbelse er både øget opløsning af befolkningsstruktur og data, der er tilstrækkelige til statistisk at udlede befolkningshistorier og teste alternative modeller for divergens (Degnan og Rosenberg, 2009; Sukumaran og viden, 2017). På den anden side afslører genomskalaanalyser også, at (som længe været anerkendt for planter), genetisk udveksling (introgression) blandt dyrearter er mere almindelig end tidligere antaget, både under og efter speciering, og kan endda drive nye adaptive strålinger (Mallet, 2007; Rieseberg og Vilis, 2007; Arnold og Kunte, 2017) Figur 1. Samlet set understreger disse indsigter yderligere, at speciering er langvarig (f.eks., 2016) og afslører de modsatte kræfter, der opererer gennem fylogeografi-fylogenetik kontinuum., 2016).
hvordan hænger alt dette sammen med taksonomi og bevaringsmål? Der er stærk og berettiget bekymring for, at den øgede opløsning, som genomskærme giver, kan føre til voldsom overspaltning af arter (Isaac et al., 2004), der potentielt begrænser ledelsesfleksibilitet og sender små genetisk divergerende populationer til indavl og eventuel udryddelse (Frankham et al., 2012). Omvendt, Gippoliti et al. (2018) hævder, at taksonomisk inerti, hvilket resulterede i tidligere manglende anerkendelse af mange arter af afrikanske hovdyr, havde resulteret i outbreeding depression efter blanding og manglende beskyttelse af geografisk begrænset taksa. Deres opfattelse (og se også Morrison et al., 2009) er, at der ikke er noget bevis for, at taksonomisk inflation har negative virkninger på bevarelse af hovdyr (Gippoliti et al., 2018). En relateret bekymring er, at ustabil taksonomi forsinker både praksis med bevarelse og offentlige opfattelser (Garnett og Christidis, 2017). Disse bekymringer er delvist drevet af den langvarige debat omkring artskoncepter og diagnostiske metoder i systematisk biologi, især anvendelse af det fylogenetiske Artskoncept, der hævder, at arter er minimalt diagnosticerbare enheder (Cracraft, 1983). Imidlertid er flertallet af taksonomer i sagens natur konservative og navngiver kun arter, når der er overensstemmelse på tværs af flere bevislinjer (Dayrat, 2005; Padial et al., 2010; Schlick-Steiner et al., 2010: Yeates et al., 2011). Vores opfattelse er, at arter skal repræsentere robuste uafhængigt udviklende slægter, der forbliver stort set intakte, når de er i kontakt med nære slægtninge—i overensstemmelse med hensigten med det evolutionære Artskoncept (1978) og det generelle Slægtskoncept (1998, 2007). I praksis kræver dette direkte bevis for reproduktiv isolering eller rimelige surrogater af det samme, samtidig med at det giver mulighed for begrænset genetisk udveksling (Singhal et al., 2018).
genomiske data giver større effekt end før til at identificere differentiering og divergens inden for og mellem arter, hvilket kræver større overvejelse af, hvordan dette påvirker afgrænsningen af taksonomiske enheder. Hvor populationer både er stærkt genetisk og fænotypisk divergerende i forhold til allerede navngivne arter, er der en stærk sag for at navngive disse som separate arter. Hvis populationer er stærkt divergerende fænotypisk med lidt genomisk adskillelse, er der en domskald på, om disse fænotypiske forskelle er (i) arvelige og (ii) sandsynligvis vil forårsage materiel reproduktiv isolation. Genomscanninger kan løse sådanne situationer ved at teste for begrænset genstrømning. Det mere udfordrende problem er, hvor morfologisk ens, men genetisk divergerende populationer opdages. Sådanne populationer kan let diagnosticeres som separate evolutionære Slægter ved at anvende statistiske afgrænsningsmetoder til multilocus-data (Carstens et al., 2013; Rannala, 2015), men er de flygtige populationer eller holdbare arter (Sukumaran og viden, 2017)? Det er bestemt muligt inden for nomenklaturens regler at genkende arter fra DNA-sekvenser alene, hvilket giver sekvensbaseret diagnostik., 2003; Cook et al., 2010). For eksempel Murphy et al. (2015) afgrænse og beskrive seks Australske desert spring amphipod arter baseret på DNA-sekvensdata på trods af ingen klare morfologiske forskelle. Dette er dog et generelt forbedringsområde, hvor systematikere er nødt til at anvende udtrykket “kryptiske arter” med større konsistens og strenghed (Struck et al., 2017).
i situationer, hvor DNA-sekvensdata giver en potentiel nøgledefinerende funktion til genkendelse af en art, foreslår vi, at det er særligt vigtigt at anvende andre beviser for at undgå oversplitting (Oliver et al., 2015). En metode er at teste for betydelig reproduktiv isolering, hvor sådanne populationer danner naturlige kontaktområder. For eksempel i en sammenlignende genomisk analyse af kontaktområder blandt afstamningspar af Carlia-firben, der er fænotypisk uadskillelige, var indekser for genetisk isolation stærkt korreleret med divergenstid (Singhal og Morits, 2013; Singhal og Bi, 2017) (figur 2). De mere divergerende af disse slægtspar er nu formelt anerkendt som separate arter, mens mere nært beslægtede (men alligevel statistisk diagnosticerbare) slægter med tegn på genomisk omfattende genetisk udveksling på tværs af kontaktområder ikke var (Singhal et al., 2018). I betragtning af det stærke empiriske forhold mellem divergens og reproduktiv isolering i disse takser adskilte disse forfattere yderligere to allopatriske og kryptiske isolater (af en nært beslægtet art), der havde endnu højere genomisk divergens end dem, der er afgrænset ovenfor. Udvidelse af denne reproduktive isolation informeret metrisk til andre kongeneriske arter med dybt divergerende, men morfologisk lignende fylogeografiske slægter (Potter et al., 2016, 2018; Afonso Silva et al., 2017a) antyder, at der stadig er flere arter, der skal beskrives, især på øer (figur 2, Carlai AMAKS og C. rufilatus), og faktisk er nogle sådanne arter blevet beskrevet efter komplementær fænotypisk analyse (Afonso Silva et al., 2017b). Andre slægter med nyere afvigelser, men stadig statistisk afgrænset som separate Slægter ved hjælp af et stort sæt nukleare gener, kan repræsenteres som evolutionært signifikante enheder (ESU ‘ er) inden for arter.
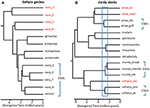
figur 2. Fylogenomisk opløsning af artsgrænser i firben fra de australske Monsoonale troper (AMT). I hvert tilfælde estimeres forhold og relative divergenstider fra 100 eksoner ved hjælp af Multispecies coalescent implementeret i StarBEAST2, men tidsskalaerne mellem de to eksempler er ikke nødvendigvis de samme. A) resultater for nana-gruppen af gehyra-gekkoer., 2018). Den fylogenetiske mangfoldighed af slægter inden for G. nana, som tidligere anerkendt, omfattede fire andre allerede beskrevne og morfologisk karakteristiske arter. Efterfølgende analyser af områdeoverlapninger og morfologi har ført til anerkendelse af hver af de mest fylogenetisk divergerende slægter (røde taksonmærker) som separate arter (Doughty et al., 2018). De (morfologisk uadskillelige) slægter, der er tilbage i G. nana som nu anerkendt, kan betragtes som ESU ‘ er. B) løbende afvikling af artsgrænser i Carlia skinks fra AMT. Nogle slægter er allerede blevet anerkendt som arter baseret på en kombination af afgrænsning af genetiske arter og post hoc-identifikation af diagnostiske morfologiske forskelle (C. insularis og C. isostriacantha; Afonso Silva et al., 2017b). For andre komplekser (Potter et al., 2016, 2018), taksonomiske revisioner er i gang. Det blå rektangel angiver rækkevidden af divergenstider, hvor der er observeret betydelig reproduktiv isolering, korreleret med divergens, i kontaktområder mellem morfologisk kryptiske slægter af beslægtede arter (Singhal og Bi, 2017). Konservativt og efterfølgende (Singhal et al., 2018), anerkender vi som kandidatarter tre genetiske slægter med forskelle større end denne empiriske tærskel. Dette ville efterlade genetisk forskellige ESU ‘ er inden for hver af Carlia AMAKS, Carlia mundaog Carlia rufilatus (blå parenteser). Skalastængerne repræsenterer estimater af divergenstid i millioner af år.
i ekstreme tilfælde afslører genomiske metoder, at enkelte taksonomiske arter ikke kun er genetisk heterogene, men i fylogenetisk forstand også indeholder andre navngivne arter. En sådan “parafyly” kan opstå gennem nylig speciering fra en enkelt, genetisk opdelt Art (Patton og Smith, 1994), men det genererer også hypotesen om, at den genetisk heterogene takson kan repræsentere flere arter. Igen kræves der flere beviser. Som et eksempel herpå afgrænsede fylogenomisk analyse af et kompleks af gehyra gecko firben fra det nordlige Australien ni evolutionære slægter, med fire allerede navngivne arter fylogenetisk indlejret blandt dem., 2018; figur 2). Efterfølgende bevis for geografisk overlapning (samtidig med at fylogenetisk adskillelse bevares) og subtile morfologiske forskelle understøttede navngivning af de fire mest fylogenetisk divergerende slægter (Doughty et al., 2018). Omvendt havde de mere nært beslægtede (men alligevel statistisk adskillelige) slægter tilstødende snarere end overlappende fordelinger, men var ikke morfologisk diagnosticerbare og blev følgelig bevaret som en enkelt art (G. nana sensu stricto), omend med flere ESU ‘ er, i afventning af yderligere undersøgelse (figur 2). Efterfølgende genomscanninger af prøver på tværs af grænserne for disse ESU ‘ er afslørede, at i det mindste nogle har betydelig genetisk udveksling, validering af originalen, konservativ tilgang taget til genkendelse af arter.i andre tilfælde kan plantegenomiske undersøgelser, selv gennem simpel ordination af SNP ‘ er, afsløre skarpe og uventede forskelle mellem populationsprøver, hvilket antyder tilstedeværelsen af kryptisk taksa, der derefter kan analyseres for diagnostiske fænotypiske tegn. For eksempel, SNP-baseret bevis for Genomisk divergens mellem populationer og økotypiske forskelle i Eucalyptus salubris afslørede to forskellige molekylære slægter, der opretholdt deres særpræg i et område med geografisk overlapning, med økotypisk tilpasning betragtes som en vigtig faktor for at minimere genstrømning mellem slægterne. (Steane et al., 2015; figur 3). I et andet tilfælde har genomisk analyse kombineret med morfologiske og reproduktive undersøgelser i Australsk Pelargonium ført til signifikante ændringer i taksonomi, både opdeling og klumpning af taksonomi, især hvor tidligere anvendte morfologiske træk viser sig at være labile og uoverensstemmende med fylogeni (Nicotra et al., 2016).
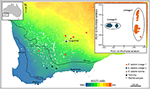
figur 3. Genomisk analyse ved hjælp af SNP-data identificerede kryptiske slægter i den udbredte Eucalyptus salubris. Placeringen af prøvepopulationer af hver slægt vises på tværs af en tørhedsindeksgradient i det sydvestlige vestlige Australien. PCoA baseret på binære genetiske afstande afledt af 16.122 dartsek markører viser to forskellige slægter. Tilpasset fra Steane et al. (2015).
ovenstående tilfælde illustrerer den progressive hypotesetestmetode, der er blevet brugt i Integrativ taksonomi (Padial et al., 2010; Yeates et al., 2011). Desuden bruger disse tilfælde en form for genomreduktionssekventering for at opnå data af høj kvalitet fra 1000-vis af uafhængige gener, hvilket giver meget større tillid til resultaterne, end det er tilfældet ved kun at bruge plastid loci eller et par nukleare gener. Et eksempel på, hvordan anvendelse af integrativ taksonomi dramatisk kan ændre opfattelsen af mangfoldighed, kommer fra en kombineret genetisk, akustisk og morfologisk undersøgelse af frøer fra Madagaskar, der næsten fordoblede den kendte artsdiversitet (Vieites et al., 2009). Disse forfattere foreslog en række udtryk, der afspejler tilstanden af taksonomisk opløsning og kongruens: Bekræftede Kandidatarter til genetisk og fænotypisk divergerende, men endnu ikke navngivet taksa; ubekræftede Kandidatarter til genetisk divergerende taksa, hvor morfologi og akustik endnu ikke var undersøgt; og dybt Konspecifikke slægter for enheder, der vides at være genetisk divergerende, men alligevel fænotypisk kryptiske. Genomiske analyser skal endnu ikke anvendes på disse takser, men ville helt sikkert informere beslutninger om specifikke slægter vs. kandidatarter.
genomiske tilgange, selv når de anvendes konservativt, vil uden tvivl føre til yderligere opdeling af morfologisk konservativ taksa. Men omvendt er der klare eksempler, hvor de samme tilgange afslører overdelte eller forkert diagnosticerede arter (Nicotra et al., 2016). Selvfølgelig er der allerede mange molekylære undersøgelser af plante-og dyrearterkomplekser baseret på datasæt i mindre skala (f. eks., mikrosatellit, mtDNA og cpDNA), der har identificeret, hvor arter er blevet fejldiagnosticeret, eller hvor flere arter er blevet omskrevet taksonomisk, men nu betragtes som en enkelt art. For eksempel Rabosky et al. (2014) brugte mitokondrie-og nukleare genfilogenier til at revidere scincid Ctenotus inornatus-komplekset, hvilket resulterede i en nettoreduktion af arter og justering af artsgrænser; de morfologiske træk, der tidligere blev anvendt, mens de var praktiske for feltbiologer, var meget labile og kunne stort set ikke matche artsgrænser som diagnosticeret genetisk. I planter findes nogle af de mest kendte eksempler i orkideerne, såsom Den Europæiske orkideslægt Ophrys og den australske orkideslægt Caladenia, hvor molekylære undersøgelser indikerer signifikant færre enheder, der ville blive anerkendt som arter, end dem, der er baseret på morfologiske træk og pseudokopulatoriske bestøvningssyndromer (se Devey et al., 2008; Svarts et al., 2014).Genomskaladata, når de fortolkes konservativt og ideelt med parallelle analyser af fænotyper, vil fortsætte med at føre til taksonomiske ændringer, især i morfologisk konservativ taksa (Fi Kerrer et al., 2018). Men i modsætning til bekymringer udtrykt af nogle (Garnett og Christidis, 2017) mener vi, at dette er den rette forfølgelse af evolutionær systematik, selvom resultaterne midlertidigt er ubelejlige.
genomik og intraspecifikke Bevaringsenheder
skønt mindre omstridt end taksonomisk afgrænsning og debatten om artsniveau, men på mange måder lige så kompleks, har været den betydningsfulde diskussion om definitionen og afgrænsningen af bevaringsenheder som vigtige elementer i intraspecifik mangfoldighed, der skal overvejes i bevaringsaktioner (Ryder, 1986; Vaples, 1991; Dison et al., 1992, 1994, Vogler og de Salle, 1994, Crandall et al., 2000; fraser og Bernatches2001; Avis, 2004; Funk et al., 2012) (figur 4). I betragtning af den forsigtige tilgang til at genkende arter fra genomiske beviser, som vi går ind for ovenfor (og se også Oliver et al., 2015; Singhal et al., 2018), er det uundgåeligt, at nogle taksonomiske arter stadig vil indeholde lange isolerede populationer i dem. Begreber som evolutionært signifikante enheder (ESU ‘er) eller Designerbare enheder (du’ er) er blevet foreslået som supplement til eksisterende taksonomi (Ryder, 1986; 1991; 1994; Mee et al., 2015). Som med arter varierer fortolkninger og kriterier, men den fælles hensigt er at genkende vigtige elementer i intraspecifik mangfoldighed for bevaringsaktioner (Funk et al., 2012). (1994, 2002) foreslog en forenklet, men praktisk molekylær tilgang til at identificere uafhængigt udviklende segmenter inden for arter, den “historiske” mangfoldighedsakse; der henviser til, at andre lægger større vægt på den “adaptive” akse for at inkludere funktionel mangfoldighed (Ryder, 1986; Crandall et al., 2000; Fraser og Bernatches2001; et al., 2001; Mee et al., 2015) (figur 4). Uanset de relative fordele ved disse alternativer er det klart, at de tekniske og analytiske værktøjer til populationsgenomik nu robust kan identificere begge komponenter i intraspecifik mangfoldighed (Funk et al., 2012; Hoban et al., 2013, 2014, Yang og Rannala, 2014).
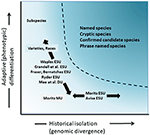
figur 4. Adskillelse af genetisk mangfoldighed i to komponenter: adaptiv variation, der opstår gennem naturlig selektion, og analyseret dog analyse af fænotyper og historisk isolation eller neutral divergens vurderet gennem genomisk divergens. Det mørkere skraverede område over kurven angiver forhold, hvor populationer sandsynligvis vil blive betragtet som separate arter under de fleste begreber. Forskellige bevaringsenhedskoncepter og infrarøde taksonomiske kategorier er vist i forhold til de to akser af genetisk mangfoldighed (efter Moritsa, 2002).
dette fører til spørgsmålet: hvorfor ikke bare navngive ESU ‘ er som underarter? Underarter er traditionelt blevet defineret og betragtet af både botanikere og dyrlæger som fænotypisk forskellige, allopatriske sæt populationer, der kan intergradere ind i hinanden ved geografiske grænser (Mayr, 1942; Grant, 1981). Som sådan afspejler de potentielt adaptiv divergens uden nødvendigvis at have været historisk isoleret. Underarter betegnelser er meget udbredt i planter, med geografiske og økologiske forskelle de primære midler, hvormed de skelnes (Hamilton og Reichard, 1992). I det omfang underarter manifesterer lokal tilpasning, med eller uden løbende genstrømning, vi foreslår, at fænotypisk definerede underarter fortsat skal anerkendes i bevaring, og ikke sammenflettet med ESU ‘er, i det mindste ikke ESU’ er som defineret til at repræsentere Historisk isolerede populationer.
på samme måde som populationsgenomik giver langt større præcision i identifikationen af ESU ‘ er ved at anvende både neutrale og adaptive loci giver det også et middel til at udnytte et stort antal neutrale loci til lettere at afgrænse styringsenheder (MUs) som demografisk uafhængige enheder karakteriseret ved begrænset genstrømning (Funk et al., 2012). Mens det meste af vores fokus her er på ESU ‘er og analoge enheder såsom Du’ er, der kan anerkendes som de største intra-specifikke enheder til bevarelse og er blevet accepteret til beskyttelse under nogle jurisdiktioner, er MUs også blevet identificeret, der repræsenterer demografisk uafhængige sæt populationer (se Palsb Larstll et al., 2007), der kan forvaltes for at bevare den større ESU (1994; Funk et al., 2012). Forvaltningsenheder kan være vigtige for overvågning og styring af populationer af arter og er også blevet omtalt som “bestande”, især relevant for fiskeriforvaltning (Avis, 2004; Laikre et al., 2005). Definitionen af MUs af Moritsa (1994) som “populationer med signifikant divergens af allelfrekvenser ved nukleare eller mitokondrie loci, uanset allelernes fylogenetiske særpræg” er generelt bredt accepteret, skønt som påpeget af Palsb Lutll et al. (2007), demografisk forbindelse er nøglefaktoren ikke niveauet for historisk genstrømning.afgrænsning af bevaringsenheder som vigtige elementer i intraspecifik mangfoldighed har også betydelige konsekvenser for genetisk redning af små indavlede populationer gennem restaurering af genstrømning eller ved forstærkning, og det bliver mere og mere klart, at genetisk redning skal overvejes bredere, hvis øget befolkningsudryddelse skal afværges., 2017; Ralls et al., 2018). Det blev foreslået af Morits (1999), at blanding af MUs, men ikke ESU ‘ er, kunne betragtes som en passende strategi for genetisk redning. Som understreget af Frankham et al. (2012) blanding af taksonomisk anerkendte arter vil generelt ikke blive betragtet som acceptabel af ledere, i hvilket tilfælde taksonomisk oversplitting kunne begrænse mulighederne for at redde små indavlede populationer. Vi foreslår, at mens en mere streng tilgang til afgrænsning af intraspecifikke enheder såsom MUs kunne hjælpe med at lindre dette problem, det er også klart, at genomisk estimering af migrationshastigheder og adaptiv divergens har potentialet til at give meget mere robust vejledning i ikke kun at definere artsgrænser og intraspecifik variation, men ved design af genetiske redningsstrategier.
Phylogenomics and Conservation
Genomics kombineret med fylogenetiske metoder kan let løse forhold og evolutionær afstand mellem taksa samt afgrænse arter. Dette opnås bedst ved hjælp af Arts træmetoder snarere end sammenkædning, da førstnævnte tegner sig for gentræ heterogenitet og undgår overinflation af spidslængder på grund af forfædres polymorfisme., 2016; Ogilvie et al., 2016). Mens massive antal loci ofte ikke er væsentlige, forbedrer inkludering af flere loci ofte opløsningen af både forhold og grenlængder (f.eks., 2016). Betydningen for bevarelse er dobbelt. Først, fylogenetiske vurderinger muliggør overvejelse af arternes eller esu ‘ ernes særpræg, ikke kun om de er beskrevet. Således EDGE-programmet (Isaac et al., 2007) går ind for vægtning af arter til bevarelse baseret på hvor meget unik evolutionær historie de repræsenterer, ud over sædvanlige IUCN-kriterier. For det andet kan grenlængder selv repræsentere biodiversitetsværdier uafhængigt af taksonomi (Faith, 1992), som har vist værdi ved prioritering af bevaringsområder (Laity et al., 2015; Rosauer et al., 2016, 2018). I denne forstand, vedtagelse af fylogenetiske målinger ved hjælp af foranstaltninger såsom fylogenetisk mangfoldighed og fylogenetisk endemisme gør det muligt at fremhæve områder med høj evolutionær mangfoldighed, der kan målrettes mod beskyttelse, og prioriteres højt i enhver planlægningsproces for forbedret bevarelse af biodiversitet. Denne tilgang kan forbedre beskyttelsen af genetisk mangfoldighed og evolutionære processer for forskellige grupper af organismer på tværs af en række geografiske skalaer (Laity et al., 2015) at træffe beslutninger om områder for at prioritere mere robust over for vagarierne om, hvordan genomisk bevis oversættes til anerkendelse af arter eller intra-specifikke enheder.
den virkelige verden—lovgivningsmæssige definitioner
Politik og lovgivningsmæssige rammer for bevarelse af biodiversitet spænder fra internationale konventioner og strategier gennem nationale strategier og lovgivning og statslig eller regional lovgivning og strategier inden for lande. De vigtigste internationale instrumenter om biologisk bevarelse omfatter konventionen om biologisk mangfoldighed, der gennemføres gennem parternes konvention (COP), Verdensarvskonventionen, den globale biodiversitetsstrategi, Montreal-processen for skovbeskyttelse, Ramsar-konventionen om vådområder, konventionen om vandrende arter, konventionen om International handel med truede arter (CITES), Den Internationale Union for bevarelse af naturen (IUCN) rødliste over truede arter og De Forenede Nationers rammekonvention om klimaændringer. Alle disse chartre legemliggør det vigtigste ved bevarelse af arter og genetisk mangfoldighed som et grundlæggende mål for bevarelse af biodiversitet, hvor arter er den enhed, der bredt er defineret til beskyttelse. Definitionerne af arter i henhold til lovgivning og internationale direktiver er imidlertid ekstremt variable. De inkorporerer taksonomisk anerkendelse af arter og intraspecifik skat, men kan også inkorporere genetiske/evolutionære begreber, der ofte er omfattet af den juridiske definition af en “art” (tabel 1).

tabel 1. Verdens lovgivning og direktiver, der definerer “arter” til notering og beskyttelse.
i betragtning af den brede vifte af infrarøde niveauer, der kan beskyttes (tabel 1), bør lovgivningen og tilknyttede listeprocesser i lande lette bevarelsen af genetisk mangfoldighed. For eksempel kan definitionen af en” art”, når man overvejer lovgivning rundt om i verden (tabel 1), omfatte: arter, underarter, sorter og geografisk og/eller genetisk forskellige populationer, mens det under Sydafrikansk lovgivning også kan omfatte Sorter, geografiske racer, stammer og hybrider. Således inkluderer en “art” under de fleste lovgivninger globalt enheder, der afspejler forskellige infrarøde niveauer, der alle er en del af en bredere artsfordeling og i varierende grad afspejler bestandsartens kontinuum. Dette er parallelt med den uundgåelige vanskelighed, som taksonomer, evolutionære biologer og bevaringsgenetikere står over for med at finde en mere konsistent og ensartet måde at specificere enheder inden for dette kontinuum.
Nogle hævder, at bekymringer vedrørende de politiske konsekvenser af arter og infrarød usikkerhed er misplaceret (Hey et al., 2003), men vi er stadig af den opfattelse, at selv om lovgivningen kan give en mekanisme til beskyttelse af genetisk mangfoldighed på en række niveauer, påhviler det biologerne at give mere klarhed og konsistens om, hvordan disse niveauer skal afgrænses fra arter til individuelle populationer. På nuværende tidspunkt står ledere og lovgivere tilbage med en forvirrende række artskoncepter og infrarøde enheder leveret af forskere, når de afgrænser enheder på tværs af bestandsartskontinuumet. For eksempel betragter alle jurisdiktioner arter, der er anerkendt af taksonomer, som nøgleenheder til bevarelse (tabel 1), men alligevel er der stadig bred debat blandt taksonomer omkring artskoncepter, og hvordan arter diagnosticeres i praksis varierer inden for og på tværs af grupper. Dette har ført til en nylig opfordring til en mere reguleret tilgang til taksonomi (Garnett og Christidis, 2017), som andre har modvirket, at regulering af taksonomi ikke er svaret (Thomson et al., 2018). Selvom vi er enige i nogle af punkterne fra Thomson et al. (2018) Vi foreslår, at det er ret naivt at foreslå, at lovgivningen selv skal ændres for eksplicit at henvise til det specifikke taksonbegreb, der er underforstået med et navn. Disse og tidligere sådanne debatter fremhæver behovet for evolutionære biologer og taksonomer at tage føringen for at sikre klarhed (gennemsigtighed) i, hvordan de oversætter genomiske og andre beviser til afgrænsning af arter eller intraspecifikke enheder, og hvordan disse taksaer vedrører relevant lovgivning for bevarelse og beskyttelse.
oversættelse til ledelse
som diskuteret ovenfor anerkender al lovgivning og direktiver artsniveauet som den grundlæggende bevaringsenhed, men giver også mulighed for anerkendelse af forskellige infrarøde enheder. Mange har også et underforstået krav til formel navngivning og taksonomisk beskrivelse af arter, når de overvejes til bevaringsprioritering og formel beskyttelse. For eksempel, under IUCN Rød notering retningslinjer notering af unavngivne arter frarådes, og der er et krav om, at den nye artsbeskrivelse vil blive offentliggjort inden for fire år efter, at arten er optaget på IUCNs Rødliste, eller den vil blive fjernet. Dette fremhæver betydningen af taksonomisk navngivning og beskrivelse af enheder på både arter og infrarøde niveauer, den begrænsede taksonomiske information for mange plante-og dyregrupper (Mace, 2004; Oliver og Lee, 2010; Vilson, 2017) og behovet for at fremskynde taksonomisk indsats, især i biodiversitetsrige områder (Riedel et al., 2013; Et Al., 2015, 2017).taksonomisk navngivne arter og intraspecifikke taksonomiske kategorier, såsom underarter og sorter, giver generelt de grundlæggende enheder til formel notering, bevaringsprioritering og beskyttelse, men i nogle jurisdiktioner anerkendes andre kategorier af intraspecifikke bevaringsenheder formelt. Disse identificeres generelt ved hjælp af kombinerede genomiske og fænotypiske data. For eksempel anerkender USA Endangered Species Act distinkte Populationssegmenter (DPSs), der bredt er baseret på ESU ‘er (1991), mens Canadian Species at Risk Act anerkender du’ er. En operationel fortolkning af” forskellige populationer ” under Canadian Species at Risk Act, dus sigter mod at identificere diskrete sæt populationer, der repræsenterer vigtige komponenter i arternes evolutionære arv, og som sandsynligvis ikke vil blive erstattet gennem naturlig spredning (Mee et al., 2015).
både du ‘er og DP’ er overlapper konceptuelt med ESU ‘ er., 2013) og er blevet anvendt mest omfattende på nordamerikanske fisk, herunder nordvest USA laksefisk (vafler, 1995) og canadiske coregonider (Mee et al., 2015) interessant nok gælder DPSs i USA kun for hvirveldyr, mens i Canada du ‘ er kan anvendes på enhver organisme, der kan opføres under dens handling., 2013), med du ‘ er, der i øjeblikket er opført for leddyr, bløddyr og planter. I Australien, ligesom Canada, kan forskellige populationer opføres for enhver organisme, der er anerkendt i henhold til Environmental Protection and Biodiversity Conservation Act 1999 (EPBC Act), selvom de ikke er en specifikt navngivet kategori, såsom DUSs og DPSs. Men til dato er de kun blevet anerkendt for hvirveldyr. I USA er der 127 hvirveldyr-DP ‘ er opført til konservering (https://ecos.fws.gov/ecp/); i Canada er der opført 70 Du ‘er, der dækker alle organismer (http://www.registrelep-sararegistry.gc.ca/species/schedules_e.cfm?id=1); og i Australien populationer af 7 hvirveldyr arter, der dækker pattedyr, fisk og fugle er blevet anerkendt som havende specifik bevaringsstatus under EPBC Act (http://www.environment.gov.au/cgi-bin/sprat/public/publicthreatenedlist.pl). Mens vurderinger af disse populationer i USA og Canada generelt bruger genetisk information., 2013), i Australien er de normalt baseret på geografisk og jurisdiktionsidentifikation, skønt de i nogle af disse tilfælde er blevet understøttet af analyse af genetisk differentiering for pattedyr (Firestone et al., 1999; Armstrong, 2006) og fish (1997; Ahonen et al., 2009).listen over infrarøde bevaringsenheder for planter og hvirvelløse dyr i Canada (DUs), men ikke i USA eller Australien, fremhæver et andet problem, der ikke specifikt er relateret til afgrænsningen af bevaringsenheder, men mere bredt til betydelige uoverensstemmelser i prioritering af bevarelse af biodiversitet; ikoniske hvirveldyr som fugle, pattedyr og fisk er i fokus i mange jurisdiktioner, mens andre organismer ikke er det. Denne præferentielle liste over populationer af visse organismer afspejler en række spørgsmål, der er mindre at gøre med videnskaben om at afgrænse bevaringsenheder og mere at gøre med listen over populationer baseret på offentlig bevidsthed, politisk hensigtsmæssighed og den karismatiske karakter af organismen, der skal listes., 2013).mens nogle populationer af kendte pattedyr, fugle og fisk i Australien er beskyttet under EPBC-loven, er der nu en betydelig mængde information baseret på molekylære undersøgelser i krybdyr, hvirvelløse dyr og planter, hvor populationer kan genkendes som ESU ‘ er eller MUs, og mange af disse er af bevaringsbetydning i betragtning af deres snævre geografiske rækkevidde, lille befolkningsstørrelse, tilbagegang og modtagelighed for truende processer. For eksempel kan slående eksempler på stærkt strukturerede populationer og dybt divergerende slægter inden for arter, der let passer til kravet om en ESU, findes i en række hvirveldyr med lav spredning fra troperne (figur 2) og andre steder. Tilsvarende i Australske hvirvelløse dyr, såsom amfipoder og tusindben, understøtter signifikant genetisk struktur og fylogeografiske mønstre afgrænsningen af diskrete befolkningsklynger som ESU ‘ er, der er geografisk forskellige, med smalle geografiske områder, begrænset til specifikke levesteder og repræsenteret af kun få populationer (Finston et al., 2007; Murphy et al., 2013; Nistelberger et al., 2014a). Ofte er disse endemiske korttrækkende endemiske skatter af bevaringsmæssig bekymring (Harvey et al., 2011) og er co-placeret i endemism hotspots (f.eks., 2017), som selv er høje prioriteter for multi-takson bevaringsopmærksomhed (Rosauer et al., 2018), såsom Kimberley-regionen og Stirling Range i det sydvestlige vestlige Australien.
i australske planter er der også et betydeligt antal populationsgenetiske og fylogeografiske undersøgelser, der identificerer forskellige befolkningsgrupper inden for en række arter fra forskellige slægter, der afspejler adskilte og historisk isolerede befolkningssystemer, geologiske og edafiske kompleksiteter og besætter kontrasterende levesteder med hensyn til vegetation og klima (se Broadhurst et al., 2017). Genetisk forskellige populationer er især tydelige hos arter fra det sydvestlige vestlige Australien (Coates, 2000; Byrne et al., 2014) med en række undersøgelser, der fremhæver signifikant genetisk struktur og stærkt divergerende slægter i en række planteslægter og familier, herunder: Acacia (Mimosaceae; Coates, 1988), Atripleks (Amaranthaceae; Shepherd et al., 2015), Banksia (Proteaceae; Coates et al., 2015), Lambertia (Proteaceae; Byrne et al., 2002), Hakea (Proteaceae; Sampson et al., 2014), Eremophila (Scrophulariaceae; Llorens et al., 2015), Pultanaea (Fabaceae; Millar og Byrne, 2013), og Calothamnus (Myrtaceae; Nistelberger et al., 2014b). I mange tilfælde anerkendes disse populationer i bevaringsplanlægning af de statslige regeringsorganer og tages i betragtning i miljøkonsekvensvurderinger (se Shepherd et al., 2015), selvom de ikke er opført til beskyttelse i henhold til lovgivningen. Vi anerkender dog, at formel anerkendelse af disse enheder vil forbedre deres bevarelse på lang sigt.
disse Australske eksempler fremhæver det stigende behov for at genkende dette lag af genetisk mangfoldighed under artsniveauet. Udfordringen er at sikre, at stærkt strukturerede befolkninger og dybt divergerende slægter, der er af bevaringsmæssig bekymring, beskyttes i lyset af konkurrerende krav om bevaringsopmærksomhed. Ud over ikoniske arter er denne udfordring også tydelig globalt med undtagelse af Canada. Manglende beskyttelse af klart afgrænsede bevaringsenheder vil sandsynligvis øge risikoen for kryptisk udryddelse og tab af betydelig genetisk mangfoldighed, som i mange tilfælde kan involvere slægter og historisk isolerede populationer, der har været inden for arter i millioner af år (2002).
konklusioner og anbefalinger
listen over arter og forskellige infra-specifikke kategorier under truet artslovgivning eller under internationale organisationer som IUCN, indikerer en forpligtelse til at beskytte genetisk mangfoldighed på tværs af artspopulationen kontinuum. Alligevel fokuserer de nuværende tilgange til bevarelse af biodiversitet stort set på artens ende af dette kontinuum med mindre opmærksomhed på genetisk mangfoldighed og infrarød variation. Dette mere begrænsede fokus på genetisk variation inden for artsniveau antyder, at mens bevarelsesledere i stigende grad anerkender værdien af eksplicit at overveje genetisk information i ledelsesaktioner og prioritering, mangler de ofte handlingsmæssig information fra forskere i biodiversitet., 2008; Laikre et al., 2010; Cook og SGR Kristian, 2017; Taylor et al., 2017). Denne kløft mellem videnskab og bevaringspraksis hindrer en mere effektiv global tilgang til bevarelse og styring af genetisk mangfoldighed.
Vi foreslår, at manglen på klarhed fra biodiversitetsforskere over bevaringsenheder i vid udstrækning er baseret på uoverensstemmelser på to niveauer. Den første vedrører flere arter begreber og de forskellige diagnostiske metoder, der anvendes til at afgrænse arter, der skaber inkonsekvens på tværs taksonomiske grupper. Det andet vedrører manglende konsistens og enighed fra taksonomer, evolutionære biologer, og bevaringsbiologer om, hvordan man behandler infra-specifik taksa og andre enheder såsom ESU ‘er, du’ er, og MUs.
mens vi mener, at lovgivningen ofte allerede er tilstrækkelig fleksibel med hensyn til, hvordan taksa (eller “arter”) defineres, argumenterer vi for, at der er behov for systematisk at adressere bestandsartskontinuumet og tydeligere definere enheder til bevarelse med større anerkendelse givet til begreber som ESU ‘ er, og hvordan disse vedrører relevant lovgivning. Vi understreger, at dette spørgsmål uden tvivl vil blive mere kritisk i betragtning af genomikens magt til at opdage differentiering i gradvist finere skalaer og dens stigende anvendelse til at afgrænse arter og infrarøde enheder.
fra den foregående analyse tilbyder vi følgende anbefalinger:
(i) genomiske data skal fortolkes konservativt ved afgrænsning af arter ved hjælp af en integrerende taksonomisk tilgang, der typisk inkluderer bevis ud over de genetiske data alene.
(ii) hvor genetikere identificerer flere slægter eller forskellige populationer inden for en art, og disse ikke berettiger til forhøjelse til artsniveau og ikke taksonomisk anerkendes, at der er stærkere drivkraft og en mere ensartet tilgang til at identificere klart berettigede intraspecifikke enheder til praktisk vurdering af bevaringsstatus eller konsekvensanalyser, og at disse udtrykkeligt anerkendes som enheder til bevarelse i forbindelse med lovgivning, der er relevant for denne jurisdiktion.
(iii) IUCN overvejer gennem sin Conservation Genetics Specialistgruppe og dens regionale undergrupper at udvikle retningslinjer for anerkendelse og mærkning af intraspecifikke enheder, der er i overensstemmelse med bedste videnskabelige praksis.
(iv) den mere nuancerede opfattelse, der kommer fra evolutionær genomik af arter og kontinuumet fra populationer til arter, bør kommunikeres mere effektivt af forskere til bevaringsledere og beslutningstagere.
mere bredt gentager vi anbefalinger, der er fremsat for nylig i en række anmeldelser (Frankham, 2010; Santamar Lyra og M, 2012; Hoban et al., 2013; Latity og Potter, 2013; Laity et al., 2015; Pierson et al., 2016; Cook og SGR prisT, 2017; Taylor et al., 2017), der fremhæver behovet for forskere til at forbedre kommunikationen og øge bevidstheden med beslutningstagere, regeringsorganer, ikke-statslige organisationer og praktikere, der er resultatet af molekylær systematik, molekylær taksonomi, fylogeografi og befolkningsgenetisk forskning kan give kritisk information om bevarelse af genetisk mangfoldighed og styring af evolutionære processer.
Forfatterbidrag
DC, MB og CM bidrog alle til forberedelsen og revisionen af manuskriptet.
interessekonflikt Erklæring
forfatterne erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
anerkendelser
dette projekt blev støttet af det australske Forskningsråd (ARC Linkage Grant No. LP120200063), Australian National University og det vestlige australske Institut for biodiversitet, bevaring og attraktioner.Afonso Silva, A. C., Bragg, J. G., Potter, S., Fernandes, C., Coelho, M. M. Og Morits, C. (2017a). Tropisk specialist versus klima generalist: diversificering og demografisk historie af søsterarter af Carlia skinks fra det nordvestlige Australien. Mol. Ecol. 26:4045–4058. doi: 10.1111 / mec.14185
CrossRef fuldtekst / Google Scholar
Afonso Silva, A. C., Santos, N., Ogilvie, H. A. og Moritsa, C. (2017b). Validering og beskrivelse af to nye nordvestlige Australske regnbue skinker med Multispecies coalescent metoder og morfologi. PeerJ 5: e3724. doi: 10.7717 / peerj.3724
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Ahonen, H. Harcourt, R. G., A. J. (2009). Nukleart og mitokondrielt DNA afslører isolering af truede grå Sygeplejerske hajpopulationer (Carcharias taurus). Mol. Ecol. 18, 4409–4421. doi: 10.1111 / j. 1365-294h.2009. 04377.P>
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Armstrong, K. N. (2006). Fylogeografisk struktur i Rhinonicteris aurantia (Chiroptera: Hipposideridae): implikationer for bevarelse. Acta Chiropterologica 8, 63-81. doi: 10.3161/1733-5329(2006)82.0.CO
CrossRef Fuld tekst / Google Scholar
Arnold, M. L., og Kunte, K. (2017). Adaptiv genetisk udveksling: en sammenfiltret historie med blanding og evolutionær innovation. Tendenser Ecol. Evol. 32, 601–611. doi: 10.1016 / j. træ.2017.05.007
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Avis, J. C. (2004). Molekylære markører, naturhistorie og Evolution. 2. udgave. Sunderland, MA; København: Københavns Universitet Press.
Google Scholar
Avis, J. C., Arnold, J., Ball, R. M., Bermingham, E., Lamb, T., Neigel, J. E., Et Al. (1987). Intraspecifik fylogeografi: mitokondrie-DNA-broen mellem populationsgenetik og systematik. Ann. Pastor Ecol. Sys. 18, 489–522. doi: 10.1146 / annurev.es.18.110187.002421
CrossRef fuldtekst | Google Scholar
Bickford, D., Lohman, D. J., Sodhi, N. S, Ng, P. K., Meier, R., blinker, K., Et Al. (2007). Kryptiske arter som et vindue på mangfoldighed og bevarelse. Tendenser Ecol. Evol. 22, 148–155. doi: 10.1016 / j. træ.2006.11.004
PubMed Abstract | CrossRef fuldtekst/Google Scholar
Blom, M. P. K., Bragg, J. G., Potter, S., C. (2016). Regnskab for usikkerhed i Gen træ estimering: sammendrag-coalescent arter træ inferens i en udfordrende stråling af australske firben. Syst. Biol. 66, 352–366. doi: 10.1093 / sysbio / sy089
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Broadhurst, L., Race, M., lav, A., Bragg, J., Catullo, R., Coates, D. J., et al. (2017). Genetisk mangfoldighed og struktur af den australske flora. Dykkere. Distribuere. 23, 41–52. doi: 10.1111 / ddi.12505
CrossRef Fuld tekst / Google Scholar
Byrne, M., Coates, D. J., Forest, F., Hopper, S. D., Krauss, S. L., Sniderman, J. M. K., Et Al. (2014). “En mangfoldig flora-art og genetiske relationer,” i plantelivet på Sandplains i det sydvestlige Australien, et globalt Biodiversitetshotspot, redaktører H. Lambers, 81-99.
PubMed Abstract
Byrne, M., Macdonald, B. Og Coates, D. J. (2002). Fylogeografiske mønstre og høj cpDNA-mangfoldighed inden for Acacia acuminata (Leguminosae:Mimosoideae). kompleks i det vestlige Australien. J. Evol. Biol. 15, 576–587. doi: 10.1046 / j. 1420-9101. 2002.00429.CrossRef fuldtekst / Google Scholar
Carstens, B. C., Pelletier, T. A., Reid, N. M. Og Satler, J. D. (2013). Hvordan man fejler ved afgrænsning af arter. Mol. Ecol. 22, 4369–4383. doi: 10.1111 / mec.12413
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Coates, D. J. (1988). Genetisk mangfoldighed og populationsgenstruktur i den sjældne Chittering græs pil, Acacia anomala domstol. Aust. J. Bot. 36:273–286. doi: 10.1071 / BT9880273
CrossRef Fuld tekst/Google Scholar
Coates, D. J. (2000). Definition af bevaringsenheder i en rig og fragmenteret flora: implikationer for styring af genetiske ressourcer og evolutionære processer i planter i det sydvestlige Australien. Aust. J. Bot. 48, 329–339. doi: 10.1071 / BT99018
CrossRef fuldtekst / Google Scholar
Coates, D. J., McArthur, S. L. og Byrne, M. (2015). Signifikant genetisk mangfoldighedstab efter patogendrevet befolkningsudryddelse i den sjældne endemiske Banksia brunnii (Proteaceae). Biol. Konservator. 192, 353–360. doi: 10.1016 / j. biocon.2015.10.013
CrossRef fuldtekst / Google Scholar
Colgan, D. J., J. R. (1997). Biokemisk genetik og anerkendelse af en vestlig bestand af den almindelige ædelfisk (Scombroidea: Gempylidae)., Australien. Ødelægge. Frisk. Res. 48, 103-118. doi: 10.1071 / MF96048
CrossRef fuldtekst / Google Scholar
konventionen om biologisk mangfoldighed (2007). Tilgængelig online på: https://www.cbd.int/doc/meetings/cop-bureau/cop-bur-2007/cop-bur-2007-10-14-en.pdf
Cook, C. N. og SGR Karrus, C. M. (2017). Tilpasning af videnskab og politik for at opnå evolutionært oplyst bevarelse. Konservator. Biol. 31, 501–512. doi: 10.1111 / cobi.12863
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
kok, L. G., Edvard, R. D., Crisp, M. D. og Hardy, N. B. (2010). Behov morfologi altid være påkrævet for nye arter beskrivelser? Inverter. Sys. 24, 322–332. doi: 10.1071 / IS10011
CrossRef Fuld tekst / Google Scholar
Cracraft, J. (1983). Artskoncepter og specieringsanalyse. Curr. Ornitol. 1, 159–187. doi: 10.1007/978-1-4615-6781-3_6
CrossRef Fuld tekst / Google Scholar
Crandall, K. A., Bininda-Emonds, O. R., Mace, G. M., R. K. (2000). Overvejer evolutionære processer i bevaringsbiologi. Tendenser Ecol. Evol. 15, 290–295. doi: 10.1016 / S0169-5347 (00) 01876-0
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Cutter, A. D. og Payseur, B. A. (2013). Genomiske signaturer for udvælgelse på linkede steder: forening af forskellen mellem arter. Nat. Pastor Genet. 14, 262–274. doi: 10.1038 / nrg3425
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Dayrat, B. (2005). Integrativ taksonomi. Biol. J. Linn. Soc. 85, 407–415. doi: 10.1111 / j. 1095-8312.2005.00503.H5
CrossRef fuldtekst / Google Scholar
de kristnes, K. (1998). “Den generelle afstamning begrebet arter, arter kriterier, og processen med speciering,” i former: arter og speciering, D. J. haard,r S. H. Berlocher, 57-75.Google Scholar (2007). Artskoncepter og artsafgrænsning. Syst. Biol. 56, 879–886. doi: 10.1080/10635150701701083
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Degnan, J. H. og Rosenberg, N. A. (2009). Gen træ uoverensstemmelse, fylogenetisk inferens og multispecies coalescent. Tendenser Ecol. Evol. 24, 332–340. doi: 10.1016 / j. træ.2009.01.009
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Devey, D. S., Bateman, R. M., Fay, M. F., J. A. (2008). Venner eller familie? Fylogenetik og artsafgrænsning i den kontroversielle europæiske orkideslægt Ophrys. Ann. Bot. 101, 385–402. doi: 10.1093 / AOB / mcm299
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Serie, A. E., Lockyer, C, Perrin, V. F., Demaster, D. P. og Sisson, J. (1992). Nytænkning af aktiekonceptet-en fylogeografisk tilgang. Konservator. Biol. 6, 24–36. doi: 10.1046 / j. 1523-1739.1992. 610024.Google Scholar
Doughty, P., Bourke, G., Tedeschi, L., Pratt, R., Oliver, P., Palmer, R., et al. (2018). Arter afgrænsning i Gehyra nana (Gekkonidae: Gekkonidae). kompleks: kryptisk og divergerende morfologisk udvikling i de australske Monsoonale troper med beskrivelsen af fire nye arter. 4403: 201-244. doi: 10.11646 / dyrepark.4403.2.1
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Dynesius, M., Og Jansson, R. (2014). Persistens af slægter inden for arter: en forsømt kontrol med specieringshastigheder. Evolution 68, 923-934. doi: 10.1111 / evo.12316
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Edvard, S. V., Potter, S., Schmitt, C. J., Bragg, J. G. og Morits, C. (2016). Retikulation, divergens og fylogeografi–fylogenetik kontinuumc. Proc. Natl. Acad. Sci. U. S. A. 113, 8025-8032. doi: 10.1073 / pnas.1601066113
CrossRef Fuld tekst/Google Scholar
tro, D. P. (1992). Bevaringsevaluering og fylogenetisk mangfoldighed. Biol. Ulemper. 61, 1–10. doi: 10.1016/0006-3207(92)91201-3
CrossRef fuldtekst / Google Scholar
Finston, T. L., Johnson, M. S., Humphreys, F. F., Eberhard, S. M. Og Halse, S. A. (2007). Kryptisk speciering i to udbredte underjordiske amfipod-slægter afspejler historiske dræningsmønstre i et gammelt landskab. Mol. Ecol. 16, 355–365. doi: 10.1111 / j. 1365-294h.2006. 03123.(1999) – Google Scholar
Firestone, K. B., Elphinstone, M. S., Shervin, V. B. Og Houlden, B. A. (1999). Phylogeographical population structure of tiger Kvolls Dasyurus maculatus (Dasyuridae: Marsupalia), en truet kødædende pungdyr. Mol. Ecol. 8, 1613–1625. doi: 10.1046 / j. 1365-294h.1999. 00745.CrossRef fuldtekst / Google Scholar
Fi Roster, C., Robinson, C. T. og Malard, F. (2018). Kryptiske arter som et vindue ind i paradigmeskiftet af artskonceptet. Mol. Ecol. 27, 613–635. doi: 10.1111 / mec.14486
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Frankham, R. (2010). Udfordringer og muligheder for genetiske tilgange til biologisk bevarelse. Biol. Konservator. 143,1919–1927. doi: 10.1016 / j. biocon.2010.05.011
CrossRef Fuld tekst/Google Scholar
Frankham, R., Ballou, J. D., Dudash, M. R., Eldridge, M. D., Fenster, C. B., Lacy, R. C., Et Al. (2012). Implikationer af forskellige artskoncepter for at bevare biodiversiteten. Biol. Konservator. 153, 25–31. doi: 10.1016 / j. biocon.2012.04.034
CrossRef fuldtekst / Google Scholar
Fraser, D. J. og Bernatchesj, L. (2001). Adaptiv evolutionær bevarelse: mod et samlet koncept til definition af bevaringsenheder. Mol. Ecol. 10, 2741–2752. doi: 10.1046 / j. 1365-294h.2001.t01-1-01411.Google Scholar
Funk, Mckay, J. K., Hohenlohe, P. A. Allendorf, F. K. (2012). Udnyttelse af genomik til afgrænsning af bevaringsenheder. Tendenser Ecol. Evol. 27, 489–496. doi: 10.1016 / j. træ.2012.05.012
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Garnett, S. T. og Christidis, L. (2017). Taksonomi anarki hæmmer bevarelsen. Natur 546, 25-27. doi: 10.1038 / 546025a
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Gippoliti, S., Cotterill, F. P. D., Sinner, D. og Groves, C. P. (2018). Virkninger af taksonomisk inerti til bevarelse af afrikansk hovdyrdiversitet: oversigt. Biol. Åb 93, 115-130. doi: 10.1111 / brv.12335
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Grant, V. (1981). Plantespeciering. København: Columbia University Press.
PubMed Abstract / Google Scholar
Hamilton, C. V. og Reichard, S. H. (1992). Nuværende praksis i brugen af underarter, sorter og forma i klassificeringen af vilde planter. Takson 41, 485-498. doi: 10.2307/1222819
CrossRef fuldtekst / Google Scholar
Harvey, M. S., Johnson, M. S., Teale, R. J., et al. (2011). Beskyttelse af uskyldige: at studere endemisk kortdistance forbedrer bevaringsresultaterne. Inverter. Syst. 25, 1–10. doi: 10.1071 / IS11011
CrossRef fuldtekst / Google Scholar
Hej, J., Vaples, R. S., Arnold, M. L., Butlin, R. K. og Harrison, R. G. (2003). Forståelse og konfrontering af artsusikkerhed i biologi og bevarelse. Tendenser Ecol. Evol. 18, 597–603. doi: 10.1016 / j. træ.2003.08.014
CrossRef fuldtekst/Google Scholar
Hoban, S. M., Hauffe, H. C., Peres-Espona, S., Arntsen, J. H., Bertorelle, G., Bryja, J., et al. (2013). At bringe genetisk mangfoldighed i spidsen for bevaringspolitik og forvaltning. Konservator. Genet. Resour. 5, 593–598. doi: 10.1007 / s12686-013-9859-y
CrossRef fuldtekst / Google Scholar
Isaac, N. J., Mallet, J. og Mace, G. M. (2004). Taksonomisk inflation: dens indflydelse på makroøkologi og bevarelse. Tendenser Ecol. Evol. 19, 464–469. doi: 10.1016 / j. træ.2004.06.004
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Isaac, N. J., Turvey, S. T., Collen, B., Vandmand, C. og Baillie, J. E. (2007). Pattedyr på kanten: bevaringsprioriteter baseret på trussel og fylogeni. PLoS en 2: e296. doi: 10.1371 / tidsskrift.pone.0000296
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Jones, M. R., og god, J. M. (2016). Målrettet fangst i evolutionær og økologisk genomik. Mol. Ecol. 25, 185–202. doi: 10.1111 / mec.13304
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Jorger, K. M., Og Schr Kristdl, M. (2013). Hvordan beskrives en kryptisk Art? Praktiske udfordringer ved molekylær taksonomi. Front. Dyr. 10:59. doi: 10.1186/1742-9994-10-59
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Laikre, L., Allendorf, F., Aroner, L. C., Baker, S., Gregovich, D. P., Hansen, M. M., et al. (2010). Forsømmelse af genetisk mangfoldighed i gennemførelsen af konventionen om biologisk mangfoldighed. Konservator. Biol. 24, 86–88. doi: 10.1111 / j. 1523-1739.2009.01425.PubMed Abstract / CrossRef fuldtekst / Google Scholar
Laikre, L., Palm, S. og Ryman, N. (2005). Genetisk populationsstruktur af fisk: konsekvenser for kystområdeforvaltning. Ambio 34.111-119. doi: 10.1579/0044-7447-34.2.111
PubMed Abstract / CrossRef Fuld tekst / Google Scholar
Laity, T., Laffan, S. V., C. E., Faith, D. P., Rosauer, D. F., Byrne, M., et al. (2015). Phylodiversity at informere bevaringspolitik: et australsk eksempel. Sci. Samlet Miljø. 534, 131–143. doi: 10.1016 / j. scitotenv.2015.04.113
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Leache, A. D. og Oaks, J. R. (2017). Nytten af enkelt nukleotidpolymorfisme (SNP). data i fylogenetik. Ann. Pastor Ecol. Syst. 48, 69–84. doi: 10.1146/annurev-ecolsys-110316-022645
CrossRef Full Text | Google Scholar
Llorens, T. M., Macdonald, B., McArthur, S., Coates, D. J., and Byrne, M. (2015). Disjunct, highly divergent genetic lineages within two rare Eremophila (Scrophulariaceae: Myoporeae). species in a biodiversity hotspot: implications for taxonomy and conservation. Biol. J. Linn. Soc. 177, 96–111. doi: 10.1111/boj.12228
CrossRef Full Text | Google Scholar
Love Stowell, S. M., Pinzone, C. A., and Martin, A. P. (2017). Overvinde barrierer for aktive interventioner for genetisk mangfoldighed. Biodivers. Konservator. 26, 1753–1765. doi: 10.1007 / s10531-017-1330-CrossRef fuldtekst / Google Scholar
Mace, G. M. (2004). Taksonomiens rolle i bevarelse af arter. Philos. Trans. R. Soc. B: Biol. Sci. 359, 711–719. doi: 10.1098 / rstb.2003.1454
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Mallet, J. (2007). Hybrid speciering. Natur 446, 279-283. doi: 10.1038 / nature05706
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Mayr, E. (1942). Systematik og Arternes Oprindelse. København: Columbia University Press.
Google Scholar
Mee, J. A., Bernatches, L., Reist, J. D., Rogers, S. M. og Taylor, E. B. (2015). Identificering af designerbare enheder til intraspecifik bevaringsprioritering: en hierarkisk tilgang anvendt på Lake hvidfisk arter kompleks (Coregonus spp.). Evol. Appl. 8, 423–441. doi: 10.1111 / eva.12247
PubMed Abstract | CrossRef Full Text | Google Scholar
Millar, M. A., and Byrne, M. (2013). Cryptic divergent lineages of Pultenaea pauciflora M.B. Scott (Fabaceae: Mirbelieae) exhibit different evolutionary history. Biol. J. Linn. Soc. 108, 871–881. doi: 10.1111/bij.12002
CrossRef Full Text | Google Scholar
Moritz, C. (1994). Defining evolutionarily significant units for conservation. Trends Ecol. Evol. 9, 373–375. doi: 10.1016/0169-5347(94)90057-4
CrossRef Full Text | PubMed Abstract
Moritz, C. (1999). Bevaringsenheder og translokationer: strategier til bevarelse af evolutionære processer. Hereditas 130, 217-228. doi: 10.1111 / j. 1601-5223.1999. 00217.CrossRef Fuld tekst | Google Scholar
CrossRef Fuld tekst / Google Scholar
CrossRef Fuld tekst / Google Scholar
CrossRef, C. (2002). Strategier til beskyttelse af biologisk mangfoldighed og de evolutionære processer, der opretholder den. Syst. Biol. 51, 238–254. doi: 10.1080/10635150252899752
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Morits, C., and Potter, S. (2013). Betydningen af et evolutionært perspektiv i bevaringspolitisk planlægning. Mol. Ecol. 22, 5969–5971. doi: 10.1111 / mec.12565
PubMed Abstract | CrossRef fuldtekst/Google Scholar
Bank, S., Bourke, G., Bragg, J. G., Doughty, P., et al. (2018). Kryptisk afstamningsdiversitet, divergens i kropsstørrelse og sympatry i et artskompleks af australske firben (Gehyra). Evolution 27, 54-66. doi: 10.1111 / evo.13380
CrossRef Fuld tekst/Google Scholar
Morrison, Lohr, J. L., Duchen, P., Vilches, R., Trujillo, D., Mair, M., et al. (2009). Virkningen af taksonomisk ændring på bevarelse: Dræber det, kan det redde, eller er det bare irrelevant? Biol. Ulemper. 142, 3201–3206. doi: 10.1016 / j. biocon.2009.07.019
CrossRef Fuld tekst / Google Scholar
Murphy, N. P., Adams, M., Gusik, M. T. og Austin, A. D. (2013). Ekstraordinær mikro-endemisme i Australske desert spring amphipods. Mol. Fylogenet. Evol. 66, 645–653. doi: 10.1016 / j. ympev.2012.10.013
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Murphy, N. P., King, R. A. og Delean, S. (2015). Arter, ESU ‘ er eller populationer? Afgrænsning og beskrivelse af morfologisk kryptisk mangfoldighed i Australske desert spring amphipods. Inverter. Syst. 29, 457–467. doi: 10.1071/IS14036
CrossRef fuldtekst / Google Scholar
Nater, A., Mattle-Greminger, M. P., Nurcahyo, A., Nuak, M. G., De Manual, M., Desai, T., et al. (2017). Morfometriske, adfærdsmæssige og genomiske beviser for en ny orangutangart. Curr. Celle Biol. 27, 3487–3498. doi: 10.1016 / j. cub.2017.09.047
CrossRef Fuld tekst / Google Scholar
Nicotra, A. B., Chong, C., Bragg, J. G., Ong, C. R., Aitken, N. C., Chuah, A., et al. (2016). Population og fylogenom nedbrydning via genotypebestemmelse-for-sekventering i Australsk Pelargonium. Mol. Ecol. 25, 2000–2014. doi: 10.1111 / mec.13584
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Nistelberger, H., Byrne, M., Coates, D. og Roberts, J. D. (2014A). Stærk fylogeografisk struktur i en tusindben indikerer Pleistocæn vicariance mellem populationer på båndede jernformationer i halvtørre Australien. PLoS en 9: e0093038. doi: 10.1371 / tidsskrift.pone.0093038
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Nistelberger, H., Gibson, N., Macdonald, B., Tapper, S.-L. og Byrne, M. (2014b). Fylogeografisk bevis for to mesiske refugier i et hotspot for biodiversitet. Arvelighed 113, 454-463. doi: 10.1038 / hdy.2014.46
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Ogilvie, H. A., Heled, J., J., D. Og Drummond, A. J. (2016). Computational ydeevne og statistisk nøjagtighed af beast og sammenligninger med andre metoder. Syst. Biol. 65, 381–396. doi: 10.1093 / sysbio / syv118
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Oliver, P. M., Keogh, J. S., C. (2015). Nye tilgange til katalogisering og forståelse af evolutionær mangfoldighed: et perspektiv fra Australsk herpetologi. Aust. J. 62, 417–430. doi: 10.1071/14091
CrossRef fuldtekst / Google Scholar
Oliver, P. M., Laver, R. J., Demello Martins, F., Pratt, R. C., Hunjan, S., og morit, C. (2017). Et nyt hotspot af hvirveldyr endemisme og et evolutionært refugium i tropisk Australien. Diver. Distribuere. 23, 53–66. doi: 10.1111 / ddi.12506
CrossRef fuldtekst/Google Scholar
Oliver, P. M. og Lee, M. S. Y. (2010). De botaniske og biologiske koder hindrer biodiversitetsforskning ved at modvirke offentliggørelse af unavngivne nye arter. Takson 59, 1201-1205. doi: 10.2307/20773990
CrossRef fuldtekst / Google Scholar
Padial, J., Miralles, A., De la Riva, I. og Vences, M. (2010). Taksonomiens integrerende fremtid. Front. Dyr. 7:16. doi: 10.1186/1742-9994-7-16
PubMed Abstract / CrossRef fuldtekst / Google Scholar
Palsb Pristll, P. J., B Pristrub prisT, M. Og Allendorf, F. H. (2007). Identifikation af ledelsesenheder ved hjælp af populationsgenetiske data. Tendenser Ecol. Evol. 22, 11–16. doi: 10.1016 / j. træ.2006.09.003
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Patton, J. L. og Smith, M. F. (1994). Parafyly, polyfyly, og arten af artsgrænser i Pocket Gophers (slægten Thomomys). Syst. Biol. 43, 11–26. doi: 10.1093 / sysbio / 43.1.11
CrossRef Fuld tekst / Google Scholar
Pierson, J. C., Coates, D. J., Oostermeijer, J. G. B., Beissinger, S. R., Bragg, J. G., Sunnucks, P., et al. (2016). Genetiske faktorer i truede arter genopretningsplaner på tre kontinenter. Front. Ecol. Environ. 14, 433–440. doi: 10.1002 / gebyr.1323
CrossRef Fuld tekst / Google Scholar
Potter, S., Bragg, J. G., Peter, B. M., Bi, K. og morit, C. (2016). Phylogenomics på tipene: udlede slægter og deres demografiske historie i en tropisk firben, Carlia AMAKS. Mol. Ecol. 25, 1367–1380. doi: 10.1111 / mec.13546
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Potter, S., Sue, A. T., Bragg, J. G., Rosauer, D. F., Roycroft, E. J. og morit, C. (2018). Pleistocæn klimatiske ændringer driver diversificering på tværs af en tropisk savanne. Mol. Ecol. 27, 520–532. doi: 10.1111 / mec.14441/fuld
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Rabosky, D. L., Hutchinson, M. N., Donnellan, S. C., Talaba, A. L. og Lovette, I. J. (2014). Fylogenetisk adskillelse af artsgrænser i en udbredt gruppe af australske skinks (Scincidae: Ctenotus). Mol. Fylogenet. Evol. 77, 71–82. doi: 10.1016 / j. ympev.2014.03.026
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Ralls, K., Ballou, J. D., Dudash, M. R., Eldridge, M. D. B., Fenster, C. B., Lacy, R. C., Et Al. (2018). Opfordring til et paradigmeskift i den genetiske styring af fragmenterede befolkninger. Konservator. LETT. 11, 1–6. doi: 10.1111 / conl.124126
CrossRef Fuld tekst / Google Scholar
Rannala, B. (2015). Kunsten og videnskaben om artsafgrænsning. Curr. Dyr. 61, 846–853. doi: 10.1093/csoolo / 61.5.846
CrossRef fuldtekst / Google Scholar
Riedel, A., Sagata, K., Suhardjono, Y. R., T Larsen, R. og Balke, M. (2013). Integrativ taksonomi på fast track-mod mere bæredygtighed inden for biodiversitetsforskning. Front. Dyr. 10:15. doi: 10.1186/1742-9994-10-15
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Rieseberg, L. H., J. H. (2007). Plantespeciering. Videnskab 317, 910-914. doi: 10.1126 / videnskab.1137729
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Rosauer, D. F., Blom, M. P. K., Bourke, G., Catalano, S., Donnellan, S., Mulder, E., Et Al. (2016). Filogeografi, hotspots og bevaringsprioriteter: et eksempel fra den øverste ende af Australien. Biol. Ulemper. 204, 83–93. doi: 10.1016 / j. biocon.2016.002
CrossRef Fuld tekst / Google Scholar
Rosauer, D. F., Byrne, M., Blom, M. P. K., Coates, D. J., Donnellan, S., Doughty, P., et al. (2018). Bevaringsplanlægning i den virkelige verden for evolutionær mangfoldighed i Kimberley, Australien, undgår usikker taksonomikonservativ. LETT. 11: e12438. doi: 10.1111 / conl.12438
CrossRef Fuld tekst / Google Scholar
Rosenblum, E. B., Sarver, B. A. J., brun, J. V., Des Roches, S., Hardvick, K. M., Hether, T. D., et al. (2012). Goldilocks møder Santa Rosalia: en flygtig specieringsmodel forklarer mønstre for diversificering på tværs af tidsskalaer. Evol. Biol. 39, 255–261. doi: 10.1007 / s11692-012-9171-Google Scholar
PubMed Abstract | CrossRef fuldtekst/Google Scholar
Rouks, C., Fra Kristian, C., Romiguier, J., Anciauks, Y., Galtier, N. og Bierne, N. (2016). Kaster lys over det grå område af speciering langs et kontinuum af genomisk divergens. PLoS Biol. 14: e2000234. doi: 10.1371 / tidsskrift.pbio.2000234
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Ryder, O. A. (1986). Art bevarelse og systematik-dilemmaet af underarter. Tendenser Ecol. Evol. 1, 9–10. doi: 10.1016/0169-5347 (86)90059-5
CrossRef fuldtekst | Google Scholar
Sampson, J. F., Byrne, M., Yates, C. J., Gibson, N., Thavornkanlapachai, R., Stankovsky, S., et al. (2014). Moderne pollen-medieret genindvandring afspejler den historiske isolering af en sjælden busk i et fragmenteret landskab. Arvelighed 112, 172-181. doi: 10.1038 / hdy.2013.89
PubMed Abstract / CrossRef fuldtekst / Google Scholar
Santamarrista, L., og M. Udvikling i biodiversitetspolitikken: nuværende huller og fremtidige behov. Evol. Appl. 5, 202–218. doi: 10.1111 / j. 1752-4571. 2011.00229.P >
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Schlick-Steiner, B. C., Steiner, F. M., Seifert, B., Stauffer, C., Christian, E. og Crosier, R. H. (2010). Integrativ taksonomi: en multisource tilgang til at udforske biodiversitet. Annu. Pastor Entomol. 55, 421–438. doi: 10.1146 / annurev-ento-112408-085432
PubMed abstrakt | CrossRef fuldtekst | Google Scholar
Seehausen, O, Takimoto, G., Roy, D. og Jokela, J. (2008). Speciering vending og biodiversitet dynamik med hybridisering i skiftende miljøer. Mol Ecol. 17, 30–44. doi: 10.1111 / j. 1365-294h.2007. 03529.PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Shepherd, K. A., Thiele, K. R., Sampson, J., Coates, D. J. og Byrne, M. (2015). En sjælden, ny art af Atripleks (Chenopodiaceae) bestående af to genetisk forskellige, men morfologisk kryptiske populationer i det tørre vestlige Australien: implikationer for taksonomi og bevarelse. Aust. Syst. Bot. 28, 234–245. doi: 10.1071 / SB15029
CrossRef fuldtekst / Google Scholar
Singhal, S., Og Bi, K. (2017). Historien rydder op messer: virkningen af tid i kørsel divergens og introgression i en tropisk suturområde. Evolution 71, 1888-1899. doi: 10.1111 / evo.13278
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Singhal, S., Hoskin, C., Couper, P., Potter, S. og morit, C. (2018). Ramme for løsning af kryptiske arter: En casestudie fra firbenene i de australske våde troper. Syst. Biol. . doi: 10.1093 / sysbio/syy
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Singhal, S., og morit, C. (2013). Reproduktiv isolering mellem fylogeografiske slægter skalaer med divergens. Proc. Roy. Soc B 280: 20132246. doi: 10.1098 / rspb.2013.2246026
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Steane, D. A., Potts, B. M., McLean, E., Collins, L., Prober, S. M., Stock, D. D., et al. (2015). Genom-dækkende scanninger afslører kryptisk befolkningsstruktur i en tør-tilpasset eukalyptus. Tree Genet. Genomer 11:33. doi: 10.1007 / s11295-015-0864-Google Scholar
CrossRef Fuld tekst/Google Scholar
Struck, T. H., Feder, J. L., Bendiksby, M., Birkeland, S., Cerca, J., Gusarov, V. I., et al. (2017). At finde evolutionære processer skjult i kryptiske arter. Tendenser Ecol. Evol. 33, 153–163. doi: 10.1016 / j. træ.2017.11.007
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Sukumaran, J., og viden, L. L. (2017). De Multispecies coalescent afgrænser struktur ikke arter. Proc. Nat. Aca. Sci. U. S. A. 114, 1607-1612. doi: 10.1073 / pnas.1607921114
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
svar, N. D., Clements, M. A., bur, C. C. og Miller, J. T. (2014). Definition af bevaringsenheder i et kompleks af morfologisk lignende, seksuelt vildledende, stærkt truede orkideer. Biol. Ulemper. 174, 55–64. doi: 10.1016 / j. biocon.2014.03.017
CrossRef fuldtekst / Google Scholar
taut, D., Arctander, P., Minelli, A., Thomas, R. H. og Vogler, A. P. (2003). Et krav om DNA-taksonomi. Tendenser Ecol. Evol. 18, 70–74. doi: 10.1016 / S0169-5347 (02)00041-1
CrossRef fuldtekst | Google Scholar
Taylor, HR. At bygge bro over kløften til bevaringsgenetik ved at identificere barrierer for implementering for bevaringsudøvere. Glob. Ecol. Konservator. 10, 231–242. doi: 10.1016 / j. gecco.2017.04.001
CrossRef fuldtekst / Google Scholar
Thomson, S. A., Pyle, R. L., Ahyong, S. T., Alonso-Alonso, M., Ammirati, J., Araya, J. F., et al. (2018). Taksonomi baseret på videnskab er nødvendig for global bevarelse. PLoS Biol. 16: e2005075. doi: 10.1371 / tidsskrift.pbio.2005075
PubMed Abstract / CrossRef fuldtekst / Google Scholar
Vieites, D. R., K. C., Andreonec, F., K. Stor undervurdering af Madagaskars biodiversitet fremgår af en integreret amfibieopgørelse. Proc. Natl. Acad. Sci. U. S. A. 106, 8267-8272. doi: 10.1073 / pnas.0810821106
CrossRef Fuld tekst
Vogler, A. P., Og de Salle, R. (1994). Diagnosticering enheder af bevarelse forvaltning. Ulemper. Biol. 8, 354–363. doi: 10.1046 / j. 1523-1739.1994. 08020354.CrossRef Fuld tekst / Google Scholar
R. S. (1991). Stillehavslaks, Oncorhynchus spp. og definitionen af” arter ” i henhold til loven om truede arter. Ødelægge. Fisk. Åb 53, 11-22.
Google Scholar
Vaples, R. S. (1995). “Evolutionært signifikante enheder og bevarelse af biologisk mangfoldighed under Loven om truede arter” i Evolution og det akvatiske økosystem: definition af unikke enheder i bevarelse af befolkningen, eds J. L. Nielsen (Bethesda, MD:American Fisheries Society), 8-27.Google Scholar, R. S., Gustafson, R. G., L. A., Myers, J. M., Jjonson, O. V., Busby, P. J., et al. (2001). Karakteriserende mangfoldighed i laks fra Stillehavet Nordvest. J. Fisk Biol. 59, 1–41. doi: 10.1111 / j. 1095-8649. 2001.tb01376.Google Scholar
CrossRef fuldtekst/Google Scholar
Vaples, R. S., Nammack, M., Cochrane, J. F. og Hutchings, J. A. (2013). En fortælling om to handlinger: truede arter notering praksis i Canada og USA. BioScience 63, 723-734. doi: 10.1093 / biovidenskab / 63.9.723
CrossRef fuldtekst / Google Scholar
Vaples, R. S., Punt, A. E. og Cope, J. M. (2008). Integration af genetiske data i forvaltningen af marine ressourcer: hvordan kan vi gøre det bedre? Fisk 9, 423-449. doi: 10.1111 / j. 1467-2979.2008. 00303.CrossRef Fuld tekst / Google Scholar
vige, J. A., Thiele, K. R., Shepherd, K. A., Butcher, R., Macfarlane, T. D. og Coates, D. J. (2015). Strategisk taksonomi i et biodiversitetslandskab: en ny tilgang til maksimering af bevaringsresultater for sjælden og dårligt kendt flora. Biodivers. Konservator. 24, 17–32. doi: 10.1007 / s10531-014-0785-4
CrossRef Fuld tekst / Google Scholar
Hvidlås, R. (2014). Forholdet mellem adaptiv og neutral genetisk mangfoldighed og økologisk struktur og funktion: en meta-analyse. J. Ecol. 102, 857–872. doi: 10.1111/1365-2745.12240
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Viley, E. O. (1978). Det evolutionære artskoncept genovervejes. Syst. Biol. 27, 17–26. doi: 10.2307/2412809
CrossRef Fuld tekst / Google Scholar
E. O. (2017). Biodiversitetsforskning kræver flere støvler på jorden. Nat. Ecol. Evol. 1, 1590–1591. doi: 10.1038 / s41559-017-0360-y
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Yang, H. H. og Rannala, B. (2014). Unguided artsafgrænsning ved hjælp af DNA-sekvensdata fra flere loci. Mol. Biol. Evol. 31, 3125–3135. doi: 10.1093 / molbev / msu279
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Yeates, D. K., Seago, A., Nelson, L., Cameron, S. L., Joseph, L. og Trueman, H. H. (2011). Integrativ taksonomi eller iterativ taksonomi? Sys. Ent. 36, 209–217. doi: 10.1111/j.1365-3113.2010.00558.x
CrossRef Full Text | Google Scholar