i begyndelsen af det nittende århundrede observerede von Reckingausen, at blodkar var dækket af et ark celler. Dette tykke lag med en celle består af endotelceller, der linjer den indre overflade af blod og lymfekar. Hos mennesker anslås det, at den samlede endotelagoverflade er omkring 35 m2, og at der er omkring 1 til 1,6 103 endotelceller. Den samlede længde af arterier, vener og kapillærer er omkring 90000 km (mere end at rejse to gange rundt om i verden). Endotelceller er flade og forbundet mellem hinanden ved hjælp af celleforbindelser. Deres vigtigste funktion er som mellemmænd mellem blodet og de andre væv og bidrager til blodegenskaber, både i normale og patologivæv. De er også vigtige for udveksling af immunceller mellem blod og væv.
1. Morfologi
endotelceller er meget fladede celler, så meget, at deres kerne er den højere struktur af cellen, selvom kernen også er fladt. Den cellulære form er tilpasset kanalen endotelcellen er foring (Figur 1, 2 og 3). I de mere smalle kapillærer kan endotelceller udvide deres cytoplasma hele blodkarets omkreds, så kanalen er en række endotelceller. I større kanaler, som arterier og vener, er mange endotelceller nødvendige for at dække kanalens samlede omkreds.
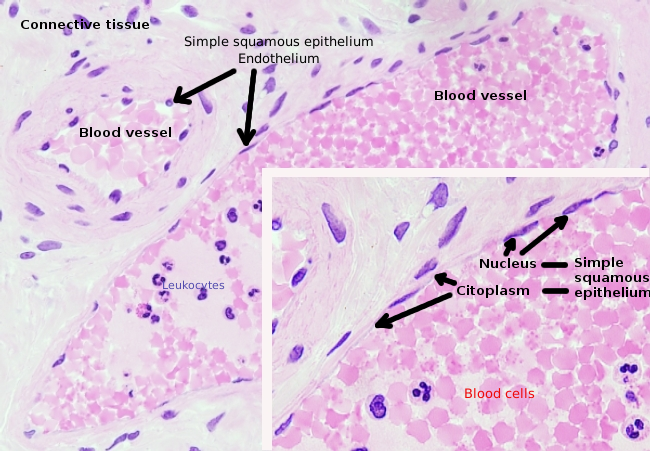
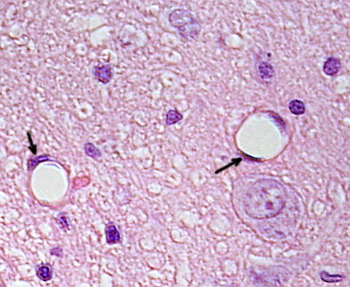
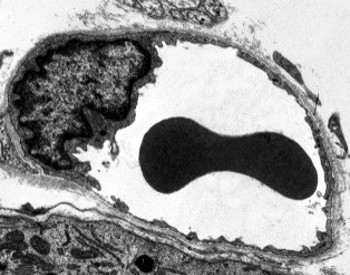
Perinuclear cytoplasma indeholder de fleste mitokondrier og andre organeller, såsom Golgi-apparater, mens perifericytoplasma er tyndere og har få organeller, skønt endoplasmatisk retikulum kan findes tæt på plasmamembranen. Endotelceller har mange vesikler, de fleste af dem er endocytiske vesikler.
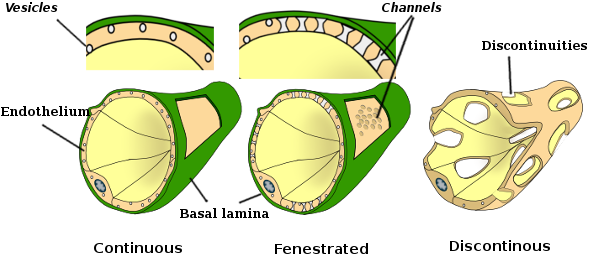
i nogle regioner, såsom lever, nyrebark og endokrine kirtler, viser endotelceller i kapillærer porer eller små passager, kendt som fenestratrioner. De er veje på omkring 60 til 70 nm i diameter, med en smal passage på omkring 5 nm, der direkte kommunikerer blod og lymfe med de andre væv. Kapillærer med disse typer endotelceller er kendt som fenestrerede kapillærer. Porerne fordeles i grupper, og deres tæthed afhænger af typen af endotel.
endotelceller adskilles fra det surronderende væv ved hjælp af et lag af ekstracellulær matrice kaldet basal lamina. Endotelcelle bidrager til dette lag med proteinerne laminin, fibronectin og kollagen type II, IV og V.
kilde og proliferation
Vaskulogenese er dannelsen af nye endotelceller i væv uden allerede eksisterende blodkar. Det sker i embryoet. Hos voksne og også i embryoner genereres nye blodkar og derfor nye endotelceller fra allerede tilstedeværende blodkar. Denne proces er kendt som angiogenese. Nye endotelceller produceres ved forgrening eller opdeling af blod og lymfekar. Angiogenese er almindelig i voksende væv eller dem under kraftig ombygning, som livmoderen hos pattedyrhunner eller i patologier som tumorer, betændelse og sår.
Vaskulogenese
første endotelceller af pattedyr genereres efter gastrulation under embryonudviklingen. De adskiller sig fra celler kaldet angioblaster, danner grupper og bliver organiseret i korte blodkar. Denne proces sker først i embryoets vitelinsæk. Disse indledende blodkar vokser og forbinder hinanden for at danne et netværk. Senere rekrutterer de fibroblaster og muskelceller. Dette indledende netværk gennemgår en kontinuerlig ombygningsproces i de næste udviklingsstadier. I embryoner er alle endotelceller oprindeligt ens, men de bliver senere differentieret i vene, arterie, kapillær og lymfekar endotel. Flere molekyler som FGF2, BMP4, IHH og VEGF inducerer adskilte differentieringsveje. Notch inducer ser ud til at være meget vigtig, fordi det fører til arterieendotel, mens dets hæmning fører til veneendotel. Lymfatisk endotel er dannet af kardinalveneendotelet og har brug for ekspression af COUPF-II og SOKS18. Derudover differentieres endotelet i overensstemmelse med det organ, det er placeret i. For eksempel danner endotel et tæt forseglet lag i hjernen for at danne blod-hjerne-barrieren, mens det er fenestreret af løst organiseret i leveren for at favorisere udvekslingen af molekyler med blodet.
da endotelceller bliver organiseret i arterie-og venekanaler, giver nogle endotelceller anledning til hæmatopoietiske celler ved en differentieringsproces kendt som endotel-hæmatopoietisk overgang (EHT). Denne proces sker også i nogle voksne væv som under dannelsen af placenta og andre væv under graviditeten.
angiogenese
angiogenese er dannelsen af nye blodkar og endotel ved forgrening eller opdeling af allerede eksisterende blodkar. Hos voksne er proliferationshastigheden af endotelceller meget lav. For eksempel kan en endotelcelle i pattedyr dele sig en gang om måneden eller vente flere år. Det betyder, at angiogenese er sjælden i væv under normal fysiologi, undtagen kvindelige reproduktive organer. Det kan dog aktiveres i patologiske processer som vævsskader og tumorer. Nogle stoffer inducerer angiogenese, såsom endotelvækstfaktor (EGF), sur EGF, fibroblastvækstfaktor (FGF), transformerende vækstfaktor (TGF) og prostaglandiner. Angiogenese begynder, når angiogene stoffer frigives fra omgivende væv. Endotelcellen i et nær blodkar, normalt fra en venule, begynder at migrere til den angiogene signalkilde og trækker de andre endotelceller, som begynder at proliferere og danne et nyt kar. Disse endotelceller mister midlertidigt deres tætte forbindelser, hvilket resulterer i afdækning af basalmembranen, som hurtigt fordøjes af endotelceller frigivet fra endotelcellerne. Nogle endotelceller går gennem den fordøjede basalmembran, mens proliferationen fortsætter. Al denne proces muliggør forlængelse og forgrening af det nye blodkar, indtil vaskulogenesesignalet forsvinder. Spidsen af blodkarrene smelter sammen med andre blodkar for at danne lukkede kredsløb og tillade strømmen af blod uden døde slutninger.
regioner med endotelcelleforfædre er fundet i dorsal aorta og endokardium. Mesenkymale celler kan differentieres fra endocardiumendotelceller og danne tricuspidventilen og nogle fibroblaster i hjertet, men ikke cardiomiocytter. Endotelceller er så fornuftige for eksterne signaler, at opretholdelse af endotelintegriteten er en aktiv proces, som medieres af andre signaler som FGF. Den aktive proces er ikke kun for integritet, men også for at bevare den cellulære fænotype. Når endotelceller ikke modtager ordentlige signaler, dør de af apoptose eller bliver mesenkymale celler, der kan syntetisere rigelig ekstracellulær matrice. Overgangen fra endotel til mesenkymal Celle ligger bag nogle sygdomme som arteriosklerose og fibrøs miokarditis.
funktion
ideen om, at endotelet kun er et passivt foringslag af lymfatiske og blodkar, skal ændres. Endotel funktioner er varierede og afgørende for organismen. Faktisk er det en fysisk struktur af kardiovaskulære og lymfekanaler, men det regulerer også udvekslingen af molekyler mellem karrets lumen og de omgivende væv, og det påvirker blodets fysiologiske egenskaber. Udover en fysisk barriere Udvikler endotelceller sekretoriske, metaboliske og immunfunktioner. De ændrer deres fysiologiske adfærd påvirket af molekyler som vækstfaktorer, koagulanter og antikoagulanter, lavdensitetslipoproteiner, serotonin, enkephalin og mange andre. Endotelceller har receptorer for alle disse stoffer.
barriere
endotelceller danner et lag, der normalt fungerer som en barriere mellem blodet og det omgivende væv. Samhørigheden mellem endotelceller opfyldes af cellekryds, såsom stramme kryds og klæbende kryds. Gap-kryds er også blevet observeret, selvom deres hovedfunktion er kommunikationen mellem tilstødende celler. Endotelceller kan modulere disse adhæsioner og ændre permeabiliteten af barrieren, der kan påvirke ikke kun til molekyler, men også til celler, der krydser endotelet. I nogle organer som lever er endotelceller imidlertid meget mere løst pakket og efterlader meget ledig plads, så endotel næppe kan betragtes som en barriere.
transmissionselektronmikroskopi billeder viser mange vesikler i cytoplasma af endotelceller. De menes at være involveret i intracellulær transport mellem den apikale (mod blodet) og basolaterale membraner (mod basal lamina). Denne type transport er kendt som transcytose. Det er interessant, at transcytosis vesikler er mere rigelige i endotelcellerne i kapillærerne end i de større blodkar. Det indikerer, at endotelcellerne i kapillærerne har en mere intens udveksling af molekyler mellem blod og væv, mens de i skibe med stor diameter hovedsageligt er involveret i at lede blodet. Nogle endotelceller, kendt som fenestreret, har porer eller meget små kanaler, der direkte forbinder blodet med det omgivende væv, hvilket tillader nogle små molekyler at krydse endotelet uden at komme ind i cytoplasmaet i nogen endotelceller (figur 4). Endelig er der organer med meget permeabelt endotel. I leveren er sinusoider blodkar, hvor endotelceller efterlader fri plads mellem hinanden, og der findes næppe en funktion som barriere.
mange celletyper rejser i blodet mod deres målorganer i kroppen. Der krydser de endotelet i blodkarrene, almindeligvis på niveau med postkapillære vener. Det betyder, at endotelceller er nødt til at modificere cellekrydsene for at lade celler gå gennem endotelaget. Leukocytter forlader blodkaret ved at genkende og forankre til specifikke molekyler af endotelcellernes apikale membran. Selectiner, integriner og immunoglobuliner er ansvarlige for genkendelse og vedhæftning af leukocytter til endotelaget. Selectiner begynder forankringen af leukocytter, som ruller over endoteloverfladen. Denne indledende vedhæftning er svag og reversibel.
mange leukocytter forlader blodet under inflammatoriske processer for at flytte til de berørte væv. Kemokiner er leukocyt-attraktive molekyler, der frigives af beskadigede væv og er forbundet med glycokalikken af endotelceller. Leukocytter, der ruller over endoteloverfladen med kemokiner, aktiveres, cellecelleadhæsion bliver stærkere, og leukocyt forbliver på stedet. Aktiveringen af leukocyten fører til aktiveringen af dets integriner, som genkender immunoglobulinerne i endotelcellen. Disse immunoglobuliner udtrykkes i endotelcellemembraner efter aktivering af cellen med kemokiner. Cellecelleadhæsionen hæver calciumkoncentrationen i endotelceller og fører til disorganisering af cellekryds og tilbagetrækning af cytoplasma. På denne måde kan leukocytter bevæge sig til grænsen af endotelceller og krydse endotelet. Adhæsionsmolekyler er også involveret i denne bevægelse af leukocytter.
blodegenskaber
endotelfunktionerne er mere komplekse end blot at kontrollere molekylerne og cellerne, der går over endotelet. Endotelceller er også involveret i blodtryk, koagulation og nogle andre blodegenskaber. primitivt kredsløbssystem menes at dukke op for 600 millioner år siden hos hvirvelløse dyr, men det manglede endotel. Endotelceller optrådte for 100 millioner år siden og gav blod en mere laminær strømning (ikke turbulent) og derfor en mere effektiv gasudveksling.
endotelceller modulerer blodtrykket ved at frigive stoffer, der virker på glat muskel i blodkar. De frigiver salpetersyre (NO) og prostacyclin, som slapper af vaskulær glat muskel. De frigiver også endothelin og blodpladeaktivatorfaktoren, hvilket begge reducerer blodkardiameteren. Nitrogenfilte frigives konstitutivt og giver en ordentlig muskeltonus, hæmmer blodpladeaggregering og leukocytadhæsion. Endothelin er en stærk vasokonstriktor. Hvordan endotelceller bestemmer, hvilket molekyle der skal frigives, er endnu ikke klart, men mekanorreceptorer i deres apikale membraner, der er i stand til at mærke blodets strømningsegenskaber, kan være plausible.
under normale forhold frigiver endotelceller molekyler i blodet, der hjælper med at opretholde en ordentlig fluiditet. De arbejder på to niveauer: fluiditet (antikoagulantia) og forebyggelse af blodpladeaggregering (antitrombotika). Proteiner C og S er vigtige molekyler, der påvirker blodets fluiditet. C-protein, der danner et kompleks med S-protein, inaktiverer koagulationsfaktorer VIIIa og Va. S protein syntetiseres af endotelceller. Endvidere indeholder endotelglycokalikset en glycosaminoglycan svarende til heparin, der er i stand til at inaktivere thrombin.
relateret til deres antikoagulerende funktion frigiver endotelceller salpetersyre og prostacyclin, hvilket begge øger den cykliske AMP i blodplader og gør vanskeligere blodpladeaggregering. Disse to molekyler frigives kontinuerligt i blodet. Endotelceller har ektonukleaser i deres apikale membraner, som fjerner ATP og ADP, begge stærke promotorer af blodpladeaggregering. Derudover frigiver endotelceller et aktivatormolekyle, der omdanner plaminogen til plasmin, hvilket favoriserer fjernelse af thrombi.
alle disse molekylære veje kan ændre sig, når endotelceller modtager nogle signaler, eller væv er beskadiget, hvilket fører til blodkoagulation og blodpladeaggregering. Under disse omstændigheder bliver endotelceller derefter aktive deltagere i koagulation og trombose.
immunforsvar
endotelceller spiller en vigtig rolle i immunforsvaret og er involveret i to mekanismer: antigenpræsentation for T-lymfocytter og rekruttering af immunceller. Sammen med makrofager kan endotelceller præsentere antigener for T-lymfocytter, fordi de konstitutivt udtrykker MHC-i (større histokompatibilitetskompleks) og kan induceres til at udtrykke MHC-II, begge nødvendige for antigenpræsentation. Endotelceller er i stand til at aktivere immunhukommelse, men ikke nye T-lymfocytter. Der er tovejsaktivering mellem endotelcelle og T-lymfocytter, så endotelcelle frigiver molekyler til at tiltrække inflammatoriske celler og udtrykker adhæsionsmolekyler til forankring af blodleukocytter.
bibliografi
Cines BD, Pollak ES, Buck CA, Loscalso J, Mcever RP, Pober JS, væge TM, Konkle BA, Schvarts BS, Barnathan ES, McCrae KR, Hug BA, Schmidt A-M, Stern DM 1998. Endotelceller i fysiologi og i patofysiologiaf vaskulære lidelser. Tidsskriftet for american society of hematology. 91:3527-3561 ![]()
Fajardo LF. 1988. Kompleksiteten af endotelceller. American journal of clinical pathology. 92:241-250.
Michiels C. 2003. Endotelcellefunktioner. Tidsskrift for cellulær fysiologi. 196: 430-443 ![]()