V časných 1960, biologové začali zkoumat, jak proteiny u různých druhů se vyvíjejí v pořadí úrovni (Zuckerkandl 1962, Margoliash 1963, Doolittle & Blombäck 1964). Proteiny analyzovány součástí hemoglobinu (Zuckerkandl & Pauling 1962), cytochrom C (Margoliash 1963), a fibrinopeptides (Doolittle & Blombäck 1964). Tyto rané výzkumy vedly k pozoruhodnému objevu: ukázalo se, že počty rozdílů mezi proteinovými sekvencemi různých druhů byly zhruba úměrné době od divergence druhů (Obrázek 1).
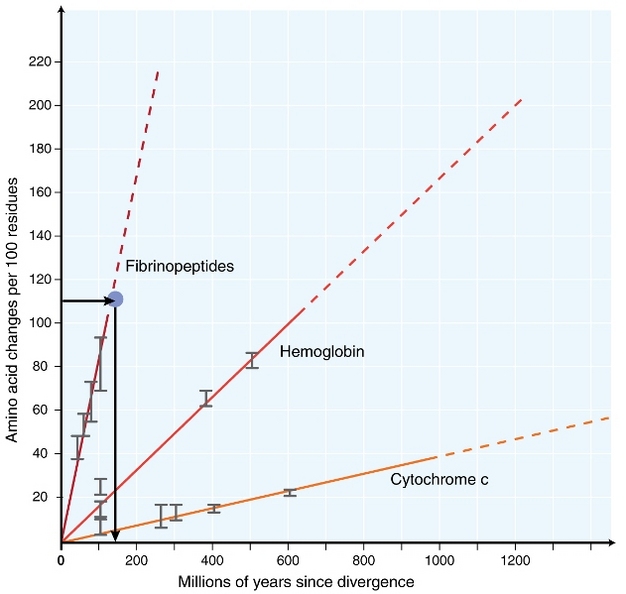
Zuckerkandl & Pauling (1965) přirovnal neustálé hromadění substituce aminokyselin v průběhu času na pravidelné ‚tikají‘ hodiny, a řekl, že tam může existovat molekulární evoluční hodiny‘. To znamená, že termín molekulární hodiny byl původně vytvořen k popisu změny v aminokyseliny vyskytující se v úměrné době od divergence druhů.
od svého prvního použití se termín „molekulární hodiny“ používá v mnoha různých kontextech. V dnešní době se často používá jednoduše odkazovat na počet změn, nebo „substituce“, nahromaděné v sekvenci DNA nebo proteinů v dané linii. Počet substitucí za definovanou jednotku času lze popsat jako „rychlost“ molekulárních hodin, což je v této souvislosti ekvivalentní „evoluční rychlosti“. Všimněte si, že počáteční konotace stále platí v některých případech. Například, když lidé říkají “ předpokládali jsme molekulární hodiny „(zejména ve fylogenetických analýzách), znamená to, že se předpokládalo, že počty substitucí se vyskytují neustále v průběhu času.
koncept konstantních molekulárních hodin má mimořádné důsledky pro evoluční biology. Pokud by skutečně existovaly konstantní molekulární hodiny, jak bylo původně navrženo, vyvození načasování evolučních událostí by se stalo poměrně přímočarým problémem (Obrázek 1). Nicméně, to stalo se jasné, že substituce se nevyskytují neustále v průběhu času v různých linií (Kumar 2005). Nicméně koncept molekulárních hodin byl mimořádně vlivný v oblasti molekulární evoluce. Jednou z nejdůležitějších myšlenek inspirovaných konceptem molekulárních hodin je neutrální teorie molekulární evoluce.
Molekulární Hodiny a Neutrality
Když myšlenka konstantní molekulární hodiny se poprvé objevil, to bylo si myslel, že převládající evoluční síly hlubších aminokyselinové nebo nukleotidové substituce byl přirozený výběr. Po této linii myšlení by konstantní molekulární hodiny naznačovaly, že adaptivní substituce u různých druhů se vyskytují neustále v průběhu času. Je však těžké vysvětlit, jak by k adaptivním substitucím došlo takovým způsobem jako hodiny. Teoreticky, osudy adaptivní mutace jsou určeny několik evolučních parametrů, jako je síla selektivní výhodu, že mutace, velikost efektivní populace, a adaptivní mutace sazby (Kimura 1983). Tyto parametry se pravděpodobně budou lišit mezi druhy, a dokonce i v rámci druhu, v závislosti na konkrétních mutacích a jejich interakcích s prostředím.
místo toho Kimura (1968, 1969) navrhl, že většina změn na molekulární úrovni má malé funkční důsledky nebo je „neutrální“. Pokud mutace nemá žádný důsledek způsobilosti, je její osud v populaci zcela určen náhodnou náhodou. To znamená, že nemůžeme předvídat, zda bude v populaci nakonec fixována konkrétní neutrální mutace. Rychlost, s jakou dochází k neutrálním substitucím v populaci, však lze předvídat, protože závisí na jediném parametru, konkrétně na míře mutace (Kimura 1968).
představme si populaci S N počtem haploidních jedinců. Pokud neutrální mutace se vyskytují v kurzu u jednoho jedince za generaci, celkový počet mutací za jednu generaci bude N-krát u. Protože všechny tyto nové mutace jsou neutrální, jejich osudy jsou zcela dána šance. Jinými slovy, všechny mutace mají stejnou šanci na dosažení fixace (což vede k „substituci“). Pravděpodobnost, že každá nová neutrální mutace dosáhne fixace, vzhledem k tomu, že došlo k substituci, je jednoduše 1 / N. Míra substituce je vypočtena jako počet nových mutací v každé generaci (Nu) násobí pravděpodobnost, že každá nová mutace dospěje fixace (1/N), což se rovná u. Jinými slovy, pro neutrální mutace, míra substituce je rovna rychlosti mutace!
Proto, pokud většina mutací jsou neutrální (jak je navrženo v neutrální teorie) a pokud mutace sazby jsou v průběhu času konstantní, substituce by mělo dojít neustále v průběhu času stejně. Pak bychom měli sledovat hodinové pravidelné míry substitucí na molekulární úrovni. Kimura (1969) tak považoval pozorování relativně konstantních molekulárních hodin v proteinových sekvencích za silnou podporu neutrální teorie molekulární evoluce.
Testování Jak Neutrální Molekulární Hodiny Běží
Podle neutrální teorie je otázka, zda nahrazování sazby jsou v průběhu času konstantní, nebo ne, je ekvivalentní, zda neutrální mutace sazby jsou v průběhu času konstantní. Z tohoto důvodu se mnoho následných studií zaměřilo na analýzu dat z neutrálních míst, aby se zjistilo, zda jsou míry neutrální mutace v průběhu času skutečně konstantní. Stručně přezkoumáme, jak jsou tyto studie implementovány, před diskusí o teoretických debatách o molekulárních hodinách. Analýzy proteinových molekulárních hodin také pokračovaly, ale debaty obklopující variace proteinových molekulárních hodin se velmi liší od debat týkajících se neutrálních molekulárních hodin, a nebudou zahrnuty v tomto článku. Zainteresovaní čtenáři by měli konzultovat Gillespie (1991), Kumar (2005), Kim and Yi (2008) a Bedford et al. (2008).
většina empirických analýz neutrálních molekulárních hodin se spoléhá na větu, že neutrální mutace lze odvodit z neutrálních substitučních sazeb (Kimura 1968, 1969). V praxi, každá studie definuje určitý typ míst v genomu jako neutrální místa, a porovnává míry substituce těchto míst mezi liniemi.
která místa v genomu jsou skutečně neutrální, nelze zcela určit, ale vědci přišli s několika užitečnými proxy. Před érou sekvenování genomu, většina dostupných sekvenčních dat byla data z sekvencí DNA kódujících proteiny. Studie často dělí sekvence DNA kódující proteiny na dva typy míst (Wu & Li 1985). První typ lokalit zahrnuje ty, u nichž by jakákoli změna vedla k substituci aminokyselin, nebo „nesynonymní místa“. Druhý typ míst, ‚synonymem stránek, včetně těch, které kódují ‚zvrhlík‘ pozice v kodonu stolu, kde změna nemá vést k aminokyselinové substituce. Například TCT a TCC kódují serin. Pokud se změní třetí pozice těchto kodonů, budou stále produkovat stejné aminokyseliny. Takové substituce by byly méně viditelné pro přirozený výběr. V důsledku toho by molekulární hodiny na synonymních místech měly být blíže neutrálním molekulárním hodinám než nesynonymní hodiny.
Jako technik sekvencování pokročilé, některé studie používají introny jako proxy pro neutrální stránky, protože nejsou začleněny do zralé mRNAs, a proto jsou více pravděpodobné, že bude neutrální (Yi et al. 2002). Sekvence neaktivní transponovatelné prvky, které byly vloženy dlouho předtím, než divergence druhů byly také často zaměstnán (tyto jsou často označovány jako ‚předků opakuje‘, např., Thomas et al., 2003). Konečně, některé studie používají nekódující sekvence DNA (sekvence po odstranění bílkovin kódování DNA sekvence) extrahované z celého genomu zarovnání na test neutrální molekulární hodiny (Elango et al. 2006, Huttley et al. 2007).
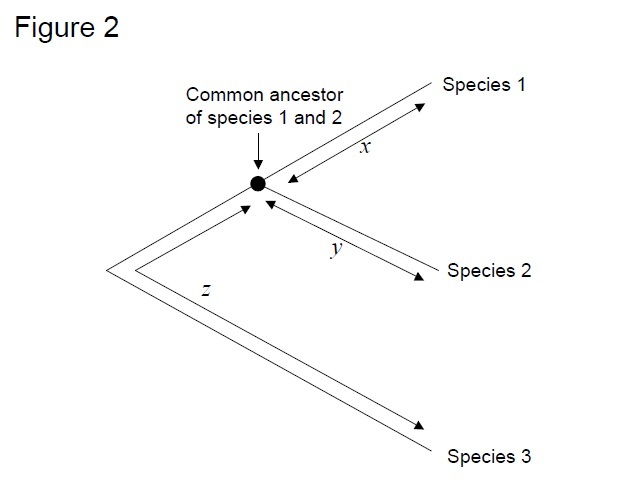
nejčastěji používaným testem je tzv. „test relativní rychlosti“ (Sarich & Wilson 1973). Zpočátku, střídání sazby za jednotku času byla odhadnuta vydělením celkového počtu rozdílů (substituce) mezi proteiny různých druhů divergence čas, odhaduje z fosilních záznamů (Obrázek 1). Fosilní záznamy však nejsou k dispozici pro mnoho srovnání a jsou spojeny s velkými chybovými okraji. Test relativní rychlosti překonává nutnost fosilních záznamů (Obrázek 2). Tak dlouho, jako outgroup sekvence na dvě linie zájmů existuje, můžeme určit, zda dvě větve sledovat stejné nebo různé molekulární hodiny, bez znalosti absolutní čas divergence (Obrázek 2).
jaké jsou determinanty neutrálních molekulárních hodin?
téměř všechny spory v srdci debat o neutrálních molekulárních hodinách vycházejí z otázky, Jaké jsou hlavní zdroje mutací. Tato otázka je přímo relevantní pro pochopení vzorců mutace, které jsou konečným zdrojem evolučních změn a genetických chorob. Dále, pochopení toho, jak se míra mutace liší mezi liniemi a genomy, je základní otázkou srovnávací genomiky, jehož cílem je použít sekvenční srovnání k identifikaci genomických oblastí, které jsou funkčně důležité.
Co tedy určuje míru neutrální mutace? Jedním z nejdůležitějších přispěvatelů k neutrálním molekulárním hodinám je změna generace specifická pro linii. Od počátku byla myšlenka konstantních neutrálních molekulárních hodin vnímána jako v rozporu s molekulárními mechanismy zárodečné mutace. To dlouho bylo za to, že většina mutací vznikají chyby v replikaci DNA v germlines (Haldane 1947, Müller 1954). Protože k mutacím dochází při replikaci zárodečné DNA pro příští generaci, měly by se hromadit v poměru k počtu generací, spíše než k absolutnímu množství času. Proto, pokud budeme srovnávat počet substitucí, které se nahromadily ve dvou linií od jejich divergence, linie s delší generační doba, které prošly méně replikace DNA událostí, by harbor méně substituce ve srovnání s linii s kratší generační doba. V důsledku toho by molekulární hodiny měly běžet pomaleji u druhů s delší dobou generace. Tato myšlenka se označuje jako „efekt generačního času“.
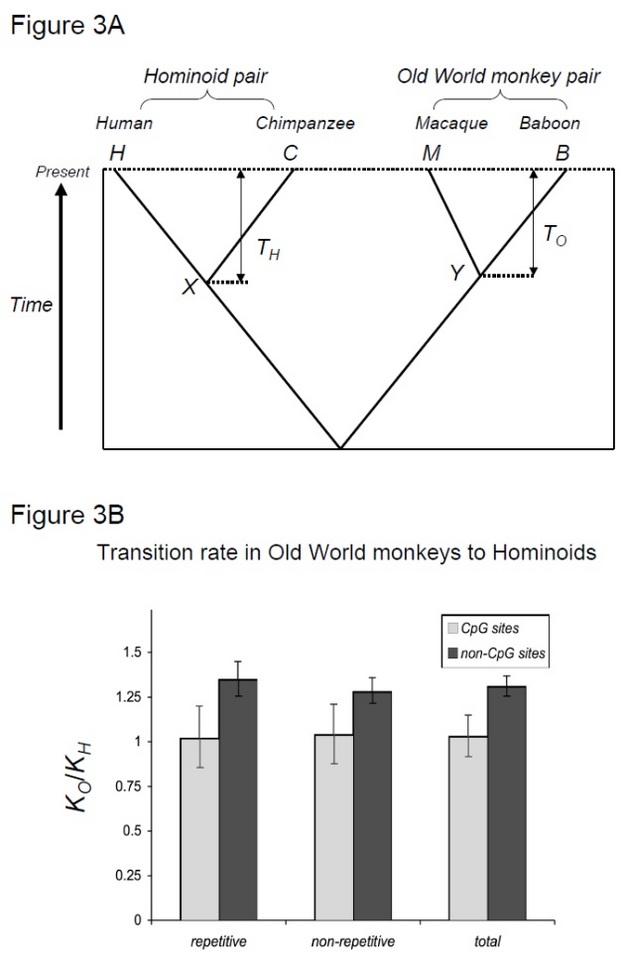
ve skutečnosti byl efekt generačního času poprvé pozorován ve studiích primátů, ještě před debatou o molekulárních hodinách. Morris Goodman, který byl pomocí imunologické metody, aby prošetřila druhů, příbuznosti v době, poznamenal, že rychlost, při které některé proteiny rozcházejí se zdá být snížena v opice, a to zejména lidé, ve srovnání s opice Starého Světa (Goodman, 1961, 1962, 1963). Tento účinek se označuje jako „zpomalení rychlosti hominoidů“. Protože hominoidy mají delší generační časy než opice Starého světa, lze toto pozorování vysvětlit efektem generačního času.
Wu & Li (1985) byli první testovací generace-time efekt hypotézy pomocí DNA sequence data. Použili data z 11 genů primátů a hlodavců. Vzhledem k tomu, že primáti mají mnohem delší dobu generace než hlodavci, molekulární hodiny by měly být u hlodavců rychlejší ve srovnání s primáty. Ve skutečnosti, zjistili, že synonymem pro místa, hlodavců ukazují, přibližně dva krát rychlost molekulární evoluce, kdy ve srovnání s primáty (Wu & Li 1985). Pro nesynonymní místa však takový účinek nebyl nalezen. Jinými slovy, neutrální molekulární hodiny, ale ne aminokyselina, molekulární hodiny tikají rychleji v hlodavec rodu ve srovnání s primáty rodu, který se hodí dobře s myšlenkou generace-time efekt.
další studie poskytla další podporu, aby hominoid rychlost zpomalení (Li & Tanimura 1987, Bailey et al. 1991) a rozdíl rychlosti mezi liniemi hlodavců a primátů (Gu & Li 1992, Huttley et al. 2007). Navíc byl pozorován rozdíl rychlosti v ještě menších fylogenetických stupnicích, zejména u primátů: například lidské molekulární hodiny běží pomaleji než šimpanzí molekulární hodiny (Elango et al . 2006); sazby u opic Nového světa jsou rychlejší než sazby u hominoidů a opic starého světa (Steiper & Young 2006). Různé rychlosti molekulárních hodin pozorované v těchto studiích jsou kvalitativně v souladu s účinkem generačního času.
skutečné rozdíly mezi liniemi však nejsou kvantitativně konzistentní s rozdílem v generačních časech. Například, Kumar & Subramanian (2002) ukázal, že i když rozdíl v generační doby mezi primátů a hlodavců je mnohem větší, než, že mezi člověka a opice Starého Světa, pozorované rozdíly v molekulární hodiny jsou podobné v těchto dvou srovnání. Je vhodné poznamenat, že Kumar & Subramanian (2002) používá specifické statistické filtry pro odstranění údajů, které ukázaly, „heterogenní“ substituční vzory, které by způsobily zkreslení směrem k pomalu se rozvíjející sekvencí (Yi et al. 2002). Nicméně, rozdíl v molekulární hodiny primátů a hlodavců se objeví mnohem méně, než původně navrhovala Wu & Li (1985). Například Huttley a kol. (2007) analyzovali kompletní genom zarovnání několika druhů včetně vačice, a ukázala, že míra rozdílu mezi eutherian linií a vačice rodu (~30%) je mnohem větší než rychlost, rozdíl mezi člověkem a myší linie (~14%). Tyto příklady ukazují, že míra rozdílů v molekulární hodiny se výrazně liší mezi různými studiemi v důsledku rozdílů v datových sadách a statistických metod. Ukazují také, že rozdíly v sazbách mezi liniemi nelze zcela zohlednit pouze rozdílem v době výroby. Jasně, existují další přispěvatelé k neutrálním molekulárním hodinám.
ve skutečnosti se zdá, že rysy historie života jiné než generace se liší s molekulárními hodinami. Martin & Palumbi (1993) ukázal, že molekulární hodiny DNA běží rychleji u druhů s malou velikostí těla. Toto pozorování vedlo k hypotéze, že metabolické rychlosti jsou důležitými determinanty molekulárních hodin. Vysoká rychlost metabolismu produkuje velké množství mutagenních kyslíkových radikálů,což by zvýšilo míru mutace (Rand 1994). Protože metabolické sazby a velikost těla obecně co-liší se generační doby, to bylo obtížné rozlišit, které z nich představuje hlavní determinantu molekulární hodiny sazby. Tsantes & Steiper (2009) navrhl, na základě údajů z primátů, že věk při první reprodukci, spíše než velikost těla, je hlavním určujícím faktorem molekulární hodiny. Vzhledem k tomu, věk při první reprodukci odráží generací čas, vliv, tato studie podporuje myšlenku, že generační doba účinku je hlavní determinantou molekulární hodiny. Tato studie je však stále založena na omezeném počtu linií (byly použity čtyři páry druhů). Tím pádem, rozlišování mezi velikostí těla, efekt generačního času, a metabolické rychlosti stále zůstávají důležitým problémem při zobecňování a porozumění neutrálním molekulárním hodinám.
dále se stále více oceňuje význam faktorů, které se nemění s dobou generace, jako je methylace DNA. Methylace DNA je chemická modifikace genomové DNA nacházející se v různých taxonech. Ve zvířecích genomech dochází k methylaci DNA téměř výhradně u cytosinů následovaných guaniny (tzv. „CpG“). Methylované cytosiny zase mají tendenci rychle mutovat na tyminy kvůli chemické nestabilitě (Bird 1980). Ve skutečnosti, v lidském genomu, mutace způsobené metylace DNA se vyskytují více než o řád častěji než jiné mutace (Nachman & Crowell 2000, Elango et al. 2008). Protože mutace způsobené methylací DNA se vyskytují do značné míry nezávisle na replikaci DNA, takové mutace mohou následovat jiné molekulární hodiny než jiné. Konkrétně, místo generace-time závislost, mutace způsobené DNA metylace může sledovat časově závislé molekulární hodiny, které je podobné tomu, co bylo původně navržené Zuckerkandl & Pauling (Kim et al. 2006)!
otestovat tuto hypotézu, Kim et al. (2006) ve srovnání člověk-šimpanz divergence na makak-pavián divergence, dva druhy páry, které sdílejí podobné divergence časy, ale s jinou generaci krát (Steiper et al. 2004). Pár člověk-šimpanz (pár hominoidů) má mnohem delší generační časy ve srovnání s párem makak-pavián (pár opic starého světa). Tato studie ukázala, že pro non-CpG míst, Staré Světové opice pár nahromaděné přibližně o 30% více substituce, což může být vysvětleno tím, že výše zmíněné hominoid míra zpomalení účinku. Naproti tomu molekulární hodiny na místech CpG vykazovaly podobný počet substitucí v párech opic hominoidů a Starého světa (obrázek 3). Tím pádem, časově závislé a časově závislé generace molekulárních hodin koexistují ve stejných genomech. Předpoklad, že pro danou linii mohou existovat jediné molekulární hodiny, již není platný, protože převládající mutační síly se mezi genomickými oblastmi liší.
Závěry
koncepce konstantní molekulární hodiny byla původně navržena na základě omezeného množství bílkovin sekvenční data. I když následné studie ukázaly, že takové pozorování není obecný vzorec aminokyselin, má významný vliv na oblasti molekulární evoluce, zejména na rozvoj neutrální teorie molekulární evoluce DNA sequence data. V návaznosti na neutrální teorii, studie se zaměřily na objasnění vzorců variací v neutrálních mutacích. Během posledních několika desetiletí jsme pozorovali, že molekulární hodiny běží mezi liniemi různými rychlostmi. Stupeň odchylky se navíc může lišit v závislosti na různých typech údajů a konkrétních použitých statistických metodách. Generace-time efekt i nadále držet na kvalitativní úrovni, ale je nedostatečné k vysvětlení kvantitativní variace neutrální mutace sazby mezi linií. Rysy historie života a mutace závislé na replikaci, jako jsou mutace způsobené methylací DNA, jsou také důležitými přispěvateli k genomickým molekulárním hodinám. Ve skutečnosti jsou různé typy molekulárních hodin pozorovány i v genomu, protože převládající mutační vstupy se liší mezi různými genomickými oblastmi. Tedy, spíše než za předpokladu, že jeden neutrální molekulární hodiny pro každý genom, aby se budoucí studie měly za cíl odhalit rozdíly genomické neutrální molekulární hodiny, aby se dozvěděli o genových mutací krajiny. Tyto informace je nejen užitečná pro pochopení surovin, jimiž se řídí molekulární evoluce a genetické onemocnění, ale také představuje rozhodující složkou ovlivňující srovnávací a funkční genomické analýzy identifikovat funkční genomové regiony.