Úvod
Mezinárodní politiky ochrany rozpozná biologické rozmanitosti na všech třech úrovních ekosystémů, druhů a genetické, a že řízení by mělo být cílem udržet všechny tři (Úmluva o Biologické Rozmanitosti, 2007). To se jasně odráží v Úmluvě o Biologické Rozmanitosti, Aichi Cíle biologické Rozmanitosti, souhlasila v roce 2010 (https://www.cbd.int/sp/targets/), tam, kde je konkrétní odkaz v cíle a cíle, a to nejen na ekosystémy a druhy, ale rovněž na genetickou rozmanitost. Přesto současné přístupy k ochraně biologické rozmanitosti jsou do značné míry založeny na zeměpisných oblastech, ekosystémů, ekologických společenstev a druhů, s menší pozornost na genetickou rozmanitost a druhy-populace kontinua. Ve skutečnosti většina z ochrany lékařů se zaměřují na druhy, jako metrické rozmanitosti, s některými úvahu vnitrodruhových jednotek v rámci intenzivního pěstování ohrožených druhů. V této souvislosti existuje přirozená touha používat jednoduché kategorizace-pojmenované druhy a další pojmenované taxony (tj. Zejména druhy poskytují společné měřítko biologické rozmanitosti, které je základem mnoha vědeckých a manažerských snah (Mace, 2004; Wilson, 2017). Jako základní jednotky zachování druhů jsou často používány kvantifikovat hodnotou biologické rozmanitosti prostřednictvím seznamy druhů v chráněných oblastech, identifikace ohrožených druhů v rámci jurisdikce, a jako základ pro biologické postupy. Kromě toho, druhy jsou obvykle entity, se kterými komunikujeme otázky ochrany veřejnosti.
tento podnět k jednoduché identifikaci diskrétních jednotek V přírodě je v rozporu s několika souvisejícími realitami. Za prvé, speciace není okamžitá. Spíše je to obvykle zdlouhavý proces (Dynesius a Jansson, 2014; Roux et al., 2016), postupuje z izolovaných populací, možná téma na jiný výběr tlaky, aby jednoznačné druhů po dlouhou dobu, váhy, s častými zvraty způsobené změnami životního prostředí nebo jednoduché genetické sloučení, jak se pohybuje připojit (viz Obrázek 1; Seehausen et al., 2008; Rosenblum a kol., 2012). Za druhé, většina uznávaných druhů není geneticky jednotná. Divergence (drift a vleklé izolace) a síťování (geneflow, hybridizace, introgression, a rekombinace) účinně tvar geografické a genomické variace v rámci druhu (Cutter a Payseur, 2013; Edwards et al., 2016) a některé druhy mohou být strukturovány do dlouho izolované populace, z nichž některé by mohly povolení uznání jako plný, ale možná morfologicky kryptických druhů (Bickford et al., 2007; Jorger a Schrödl, 2013). Za třetí, jak dlouho známo, pojmenované druhy nejsou rovnocenné v tom smyslu, kolik evoluční dědictví, které zastupují—jednoduše řečeno, ptakopysk (jako zástupce dávných dobách lišily linie) může být považována za víc, než v poslední době rozcházely hlodavce (Isaac et al., 2007). Začtvrté, označení druhů nejsou vržena do kamene, spíše se jedná o hypotézy, které mají být testovány, jakmile budou k dispozici nové důkazy (Hej et al., 2003). Každá z těchto otázek vytváří složitost a nejednoznačnost při vymezení druhů obecně, a přiřazení ochrany pozornosti k nim nebo jejich populacím. Kromě toho vymezení druhů podléhá také různým druhovým koncepcím a způsobům jejich diagnostiky, což způsobilo velké zděšení mezi biology ochrany (Mace, 2004; Frankham et al., 2012; Garnett a Christidis, 2017).
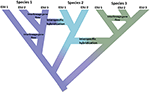
Obrázek 1. Speciace jako kontinuum postupující od izolovaných populací k jednoznačným druhům, které se projevují posunem barvy s barvami představujícími různé druhy. Některé linie, které vznikají v důsledku populačního izolace rozvinout do skutečné druhů s vnitřní uzly stromu představuje zahájení procesu speciace. Jiné linie se nevyvíjejí ve skutečné druhy, ale v průběhu času se liší, což naznačuje, že druhy nejsou geneticky jednotné. Tyto linie jsou zobrazeny jako evolučně významné jednotky (esu). Tok genů mezi liniemi je indikován pro druhy 1 A Druh 3. V tomto případě může být evoluční nezávislost přechodná, protože dvě linie patřící ke stejnému druhu nejsou reprodukčně izolovány. I když zůstávají jako odlišné linie (druhy), mezidruhová hybridizace může nastat s omezenou výměnou genů (posunem barvy) mezi druhy.
Provádění účinných strategií ochrany přírody, jejichž cílem je ochrana obou druhů a genetické rozmanitosti by měly prospěch z více konzistentní pohled na to, jak budeme vymezovat druhy a jiných jednotek v celé populaci-druhy kontinua, a jak tyto jednotky jsou interpretovány v různých právních předpisů po celém světě. Nové schopnosti z genomiky poskytují příležitosti i výzvy k dosažení těchto cílů. V následujícím textu uvažujeme: (i) jak genomu-rozsah analýzy může být v souladu s konzervativní přístup k vymezení a pojmenování druhů, a zároveň určení hlavních složek genetická rozmanitost v rámci druhů, (ii) jak stávajících právních předpisů na ochranu přírody napříč několika kontinenty, které definuje „druhů“, aby zahrnovala široké spektrum infra-zvláštní kategorie, může být použit s ohledem na vysoké rozlišení nabízených genomu obrazovky, a (iii) jak vhled do speciace procesy a strukturování odchylka od genomika sladit s právními předpisy a pak se do vedení. Poté navrhujeme několik způsobů, jak postupovat při sladění ochranářských jednotek s legislativními přístupy, abychom chránili celé spektrum rozmanitosti od populací přes druhy.
Genomika, Speciace a Taxonomie
stejně Jako mnoho oborů biologie, systematika, praxe pojmenování taxonů, a pochopení jejich vztahy a jak se tvoří, je výrazně pokročilá tím, genomika a statistické modely. Ať už prostřednictvím sekvenování celého genomu (např., 2017) nebo vzorkování genomu (SNPs, cíl capture-sekvenování; Jones a Good, 2016; Leache a Oaks, 2017), nyní máme bezprecedentní řešení vzorců genetické rozmanitosti (Funk et al ., 2012). Ve spojení se stále silnějšími statistickými modely pro odvozování historie genomové a druhové divergence, tato data poskytují nový pohled na evoluční procesy, které generují druhy a genetické variace.
aplikace těchto nových nástrojů generuje dva protichůdné poznatky. Na jedné straně mnoho studií odhaluje vysoce odlišné genetické populace v rámci pojmenovaných druhů, tzv., 2007; Jorger a Schrödl, 2013; Struck et al., 2017). To není nic nového, protože navazuje na dlouhou historii prostorové populační genetiky a molekulární fylogeografie, která byla produktivní snahou po tři desetiletí (Avise et al ., 1987). Co genomika přináší do tohoto úsilí je zvýšení rozlišení struktury obyvatelstva a dat, které jsou dostatečné k statisticky odvodit, populace, historie a vyzkoušet alternativní modely divergence (Degnan a Rosenberg, 2009; Sukumaran a Knowles, 2017). Na druhou stranu, genom měřítku analýzy jsou také ukázat, že (jak dlouho uznávána pro rostliny), genetické výměny (introgression) mezi druhy zvířat je častější, než se dříve myslelo, a to jak během a po speciace, a může dokonce řídit nové adaptivní záření (Mallet, 2007; Rieseberg and Willis, 2007; Arnold a Kunte, 2017) Obrázek 1. Dohromady tyto poznatky dále zdůrazňují, že speciace je zdlouhavá (Roux et al., 2016) a odhalit protichůdné síly působící prostřednictvím kontinua fylogeografie-fylogenetiky (Edwards et al., 2016).
Jak to všechno souvisí s taxonomií a cíli ochrany přírody? Tam je silné a odůvodněné obavy, že zvýšené rozlišení poskytované genomu obrazovky by mohlo vést k nekontrolovatelné nad-rozdělení druhů (Isaac et al., 2004), potenciálně omezující flexibilitu řízení a odesílání malých geneticky odlišných populací k inbreedingu a eventuálnímu vyhynutí (Frankham et al ., 2012). Naopak, Gippoliti et al. (2018) tvrdí, že taxonomické setrvačnost, což v předchozí selhání rozpoznat mnoho druhů Afrických kopytníků, měl za následek outbreeding deprese po míchání a neschopnost chránit geograficky omezené taxonů. Jejich pohled (a viz také Morrison et al., 2009) je to, že neexistuje žádný důkaz, že taxonomická inflace má negativní vliv na ochranu kopytníků (Gippoliti et al., 2018). Související obavou je, že nestabilní taxonomie zpomaluje jak praxi ochrany, tak vnímání veřejnosti (Garnett a Christidis, 2017). Tyto obavy jsou částečně poháněny dlouho běží debata kolem druhy, pojmy a diagnostické metody v systematické biologie, a to zejména použití Fylogenetických Druhů Koncept, který má za to, že druhy jsou minimálně rozpoznatelný jednotek (Cracraft, 1983). Nicméně, většina taxonomové jsou ze své podstaty konzervativní, pouze pojmenování druhů, kdy je shoda mezi více řádky důkazů (Dayrat, 2005; Padial et al., 2010; Schlick-Steiner et al., 2010: Yeates et al., 2011). Náš názor je, že druhy by měly představovat robustní samostatně se vyvíjející linie, které zůstávají do značné míry neporušený, když v kontaktu s blízkými příbuznými—v souladu se záměrem Evoluční Druh Koncept (Wiley, 1978) a Generalizované Linie Druhů Koncepce (de Queiroz, 1998, 2007). V praxi to vyžaduje přímý důkaz o reprodukční izolaci nebo jejich přiměřených náhradách, přičemž umožňuje omezenou genetickou výměnu (Singhal et al ., 2018).
genomická data poskytují větší sílu než dříve k identifikaci diferenciace a divergence uvnitř a mezi druhy, což vyžaduje větší zvážení toho, jak to ovlivňuje vymezení taxonomických jednotek. Kde populace jsou oba silně geneticky a fenotypově odlišné vzhledem k již vyjmenovaných druhů, existují silné argumenty pro pojmenování těchto jako samostatné druhy. Pokud populace jsou výrazně odlišná podle fenotypových znaků, s malou genomické oddělení, pak je rozhodnutí o tom, zda tyto fenotypové rozdíly jsou (i) dědičné, a (ii) mohou způsobit podstatné reprodukční izolace. Skenování genomu může takové situace vyřešit testováním omezeného toku genů. Náročnější otázkou je, kde jsou morfologicky podobné, ale geneticky odlišné populace detekovány. Takové populace mohou být snadno diagnostikovány jako samostatné evoluční linie použitím statistických metod vymezení na multilocus data (Carstens et al ., 2013; Rannala, 2015), ale jsou to pomíjivé populace nebo trvanlivé druhy (Sukumaran a Knowles, 2017)? Určitě je možné v rámci pravidel nomenklatury rozpoznat druhy pouze ze sekvencí DNA a poskytnout sekvenční diagnostiku (Tautz et al ., 2003; Cook et al., 2010). Například Murphy et al. (2015) vymezit a popsat šest australských pouštních jarních druhů amphipod na základě údajů o sekvenci DNA navzdory žádným jasným morfologickým rozdílům. Jedná se však o obecnou oblast pro zlepšení, kde systematici potřebují používat termín „kryptické druhy“ s větší konzistencí a přísností (Struck et al., 2017).
V situacích, kdy je DNA sekvence data poskytuje potenciální klíč definující funkce pro rozpoznávání druhu, jsme naznačují, že to je obzvláště důležité použít další řádky důkazů, aby se zabránilo oversplitting (Oliver et al., 2015). Jedním z přístupů je testování podstatné reprodukční izolace, pokud takové populace tvoří přirozené kontaktní zóny. Například, v komparativní genomová analýza kontaktních zón mezi lineage-dvojici Carlia ještěrky, které jsou fenotypově k nerozeznání, indexy genetické izolaci byly silně korelovány s rozdíly času (Singhal a Moritz, 2013; Singhal a Bi, 2017) (Obrázek 2). Více odlišné z těchto řádků-pair mají nyní byl formálně uznán jako samostatný druh, vzhledem k tomu, více úzce souvisí (zatím statisticky rozpoznatelný) linií s důkazy o genomically rozsáhlou výměnu genetické informace přes kontaktní zóny nebyly (Singhal et al., 2018). Vzhledem k silné empirický vztah mezi divergencí a reprodukční izolace v těchto taxonů, tito autoři dále odděleny dvě allopatric a záhadné izolátů (blízce příbuzné druhy), které měly i vyšší genomová divergence, než těch, vymezených výše. Rozšíření této reprodukční izolace informován, metrické jiných podobných skupin druhů s hluboce rozdílná, ale morfologicky podobné phylogeographic linií (Potter et al., 2016, 2018; Afonso Silva et al., 2017a) naznačuje, že je třeba popsat více druhů, zejména na ostrovech, (Obrázek 2, Carlai amax A C. rufilatus), a skutečně, některé takové druhy byly popsány po komplementární fenotypové analýze (Afonso Silva et al., 2017b). Další linií s více nedávné rozdíly, ale stále statisticky vymezené jako samostatné linie pomocí rozsáhlé sady jaderných genů, může být reprezentován jako Evolučně Významné Jednotky (ESUs) v rámci druhů.
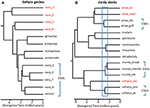
Obrázek 2. Fylogenomické rozlišení hranic druhů u ještěrů z australských monzunových tropů (AMT). V každém případě vztahy a relativní divergence době se odhaduje od 100 exons pomocí vícedruhových splývající realizován v StarBEAST2, ale čas váhy mezi dva příklady nejsou nutně stejné. A) výsledky pro skupinu Gehyra gekonů nana (Moritz et al., 2018). Fylogenetická rozmanitost linií v rámci G. nana, jak bylo dříve uznáno, zahrnovala čtyři další již popsané a morfologicky charakteristické druhy. Následné analýzy rozsah překrývání a morfologie vedlo k uznání každého z nejvíce fylogeneticky odlišných linií (červená taxonu štítky) jako samostatný druh (Doughty et al., 2018). (Morfologicky neoddělitelné) linie zbývající v G. nana, jak je nyní uznáváno, lze považovat za ESUs. (B) pokračující řešení hranic druhů v carlia skinks z AMT. Některé linie již byly uznány jako druhy na základě kombinace vymezení genetických druhů a post hoc identifikace diagnostických morfologických rozdílů (C. insularis A C. isostriacantha; Afonso Silva et al., 2017b). Pro jiné komplexy (Potter et al., 2016, 2018), probíhají taxonomické revize. Modrý obdélník označuje rozsah divergence časy, kdy značnou reprodukční izolace, souvisí s rozdíly, které byly pozorovány v kontaktních zónách mezi morfologicky kryptických linií příbuzných druhů (Singhal a Bi, 2017). Konzervativně a následovně (Singhal et al., 2018), uznáváme jako kandidátský druh tři genetické linie s odchylkami většími než tento empirický práh. To by se nechat geneticky odlišných Ježíši v každé z Carlia amax, Carlia mundo, a Carlia rufilatus (modrá závorkách). Stupnice představují odhady doby divergence v milionech let.
V extrémních případech, genomické metody odhalit jediný taxonomický druh, aby být nejen geneticky heterogenní, ale v fylogenetický smysl, aby také obsahovat další vyjmenované druhy. Takové „paraphyly“ mohou vzniknout, i když nedávné speciace z jednoho, geneticky rozdělena druhů (Patton a Smith, 1994), ale také vytváří hypotézy, že geneticky heterogenní taxon může představovat více druhů. Opět je zapotřebí více důkazů. Jako případ v bodě, phylogenomic analýzu komplexu Gehyra gecko ještěrky ze severní Austrálie oddělený devět evolučních linií, se čtyřmi již vyjmenovaných druhů fylogeneticky vložené mezi ně (Moritz et al., 2018; Obrázek 2). Následné důkazy o zeměpisné překrývání (při zachování fylogenetický oddělení) a jemné morfologické rozdíly podporované pojmenování čtyř nejvíce fylogeneticky odlišných linií (Doughty et al., 2018). Naopak, více úzce souvisí (zatím statisticky oddělitelné) linie měla přilehlých spíše než překrývající se distribucí, ale nebyly morfologicky rozpoznatelný, a proto byly zachovány jako jeden druh (G. nana sensu stricto), i když s více ESUs, až do dalšího šetření (Obrázek 2). Následné genomu skenování vzorků přes hranice těchto Ježíš ukázal, že alespoň některé mají značné genetické výměny, ověření původního, konzervativního přístupu k rozpoznání druhů.
V jiných případech, rostlina genomické studie, i přes jednoduché svěcení Snp, může odhalit stark, a neočekávané rozdíly mezi vzorky populace, což naznačuje přítomnost kryptických taxonů, které pak mohou být analyzovány pro diagnostické fenotypové znaky. Například, SNP, na základě důkazů pro genomová divergence mezi populacemi a ecotypic rozdíly v Eukalyptus salubris odhalila dva odlišné molekulární linií, které udržuje jejich rozlišovací způsobilosti v oblasti geografických překrývají, s ecotypic adaptace považována za důležitý faktor v minimalizaci toku genů mezi druhy. (Steane et al., 2015; obrázek 3). V jiném případě, v Australské Pelargonium, genomické analýzy, v kombinaci s morfologická a reprodukční studie, vedla k významným změnám v taxonomii, jak rozdělit a házet je do taxonomických jednotek, zejména tam, kde dříve používané morfologické znaky jsou labilní a nesouhlasný s fylogeneze (Nicotra et al., 2016).
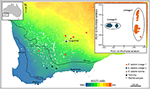
obrázek 3. Genomická analýza pomocí dat SNP identifikovala kryptické linie v rozšířeném eukalyptu salubris. Umístění vzorků populace každé linie jsou zobrazeny napříč gradientem indexu suchosti v jihozápadní západní Austrálii. PCoA založená na binárních genetických vzdálenostech odvozených od 16 122 markerů DArTseq ukazuje dvě odlišné linie. Převzato z Steane et al. (2015).
výše uvedené případy ilustrují progresivní přístup testování hypotéz, který byl použit v integrační taxonomii (Padial et al., 2010; Yeates et al., 2011). Dále, tyto případy použít nějakou formu genomu snížení sekvenování získat vysoce kvalitní data z 1000 nezávislých genů, dává mnohem větší důvěru ve výsledky, než je tomu v případě použití jen plastid loci nebo několika jaderných genů. Příklad toho, jak uplatňovat integrativní taxonomie může dramaticky změnit vnímání rozmanitosti pochází z kombinované genetické, zvukové a morfologické průzkum žab z Madagaskaru, který téměř zdvojnásobil známo, že druhová rozmanitost (Vieites et al., 2009). Tito autoři navrhli řadu pojmů odrážejících stav taxonomického rozlišení a shody: Potvrdil Kandidát Druhů, geneticky a fenotypově odlišné, ale zatím není pojmenované taxony; Neoficiální Kandidát Druhů pro geneticky odlišné taxony, kde morfologie a akustika dosud nebyly zkoumány, a Hluboce Conspecific Linií pro subjekty známo, že být geneticky odlišné, přesto fenotypově záhadné. Genomické analýzy se na tyto taxony teprve použijí, ale určitě by informoval o rozhodnutích o konspecifických liniích vs. kandidátských druzích.
genomické přístupy, i když jsou aplikovány konzervativně, nepochybně povedou k dalšímu rozdělení morfologicky konzervativních taxonů. Ale naopak existují jasné příklady, kdy stejné přístupy odhalují nadměrně rozdělené nebo nesprávně diagnostikované druhy (Nicotra et al., 2016). Samozřejmě již existuje mnoho molekulárních studií komplexů rostlinných a živočišných druhů založených na datových sadách menšího rozsahu (např., mikrosatelitů, mtDNA a cpDNA), které byly identifikovány druhy byly misdiagnosed, nebo tam, kde více druhy byly taxonomicky ohraničený, ale jsou nyní považovány za jeden druh. Například Rabosky a kol. (2014) použili mitochondriálních a jaderných genů phylogenies revidovat scincid Ctenotus inornatus komplexu, což má za následek čisté snížení druhů a přeskupení hranice druhů, morfologické znaky, aplikované dříve, zatímco vhodné pro terénní biology, byly vysoce nestabilní a do značné míry nepodařilo, aby odpovídaly hranice druhů jako diagnostikovaných geneticky. V rostlinách, některé z nejvíce dobře známé příklady jsou nalezené v orchideje, jako jsou Evropské orchideje rodu Ophrys a Australské orchideje rodu Caladenia, kde molekulární studie ukazují, výrazně méně subjektů, které by byly považovány za druhy, než ty, na základě morfologických a pseudocopulatory opylovací syndromy (viz Devey et al., 2008; Swarts et al., 2014).
data z genomu, pokud budou interpretována konzervativně a ideálně s paralelními analýzami fenotypů, budou i nadále vést k taxonomickým změnám, zejména u morfologicky konzervativních taxonů (Fišer et al ., 2018). Ale, na rozdíl od obav, které vyjádřili někteří (Garnett a Christidis, 2017), domníváme se, že toto je správná snaha o evoluční systematika, i když výsledky jsou dočasně nepohodlné.
Genomika a Zachování Vnitrodruhové Jednotky
i když méně sporné, než taxonomické vymezení a druhy úroveň diskuse, ale v mnoha způsoby, stejně jako komplex, byl významný diskuse o definici a vymezení ochrany jednotky jako důležité prvky vnitrodruhové rozmanitosti, že je třeba věnovat pozornost v ochraně akce (Ryder, 1986; Waples, 1991; Dizon et al., 1992; Moritz, 1994; Vogler a de Salle, 1994; Crandall et al., 2000; Fraser and Bernatchez, 2001; Avise, 2004; Funk et al., 2012) (obrázek 4). Vzhledem k opatrnému přístupu k rozpoznávání druhů z genomických důkazů, které obhajujeme výše (a viz také Oliver et al., 2015; Singhal a kol., 2018), je nevyhnutelné, že některé taxonomické druhy v nich budou stále obsahovat dlouho izolované populace. Pojmy jako Evolučně Významné Jednotky (ESUs) nebo Designatable Jednotek (DUs) byly navrženy, aby doplňovaly stávající taxonomie (Ryder, 1986; Waples, 1991; Moritz, 1994; Mee et al., 2015). Stejně jako u druhů se interpretace a kritéria liší, ale společným záměrem je rozpoznat hlavní prvky intraspecifické rozmanitosti pro ochranářské akce (Funk et al., 2012). Moritz (1994, 2002) obhajoval zjednodušující, ale praktický molekulární přístup k identifikaci nezávisle se vyvíjejících segmentů v rámci druhů, „historické“ osy rozmanitosti; vzhledem k tomu, že jiní kladou větší důraz na „adaptivní“ osy zahrnují funkční rozmanitosti (Ryder, 1986; Crandall et al., 2000; Fraser and Bernatchez, 2001; Waples et al., 2001; Mee et al., 2015) (obrázek 4). Ať už relativní přínos těchto alternativ, je jasné, že technické a analytické nástroje populační genomika nyní mohou spolehlivě identifikovat obě složky vnitrodruhové rozmanitosti (Funk et al., 2012; Hoban a kol., 2013; Whitlock, 2014; Yang a Rannala, 2014).
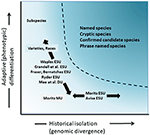
obrázek 4. Oddělení genetické rozmanitosti do dvou složek: adaptivní variace, která vzniká přirozeným výběrem, a testováno analýzou fenotypů a historickou izolací nebo neutrální divergencí hodnocenou genomickou divergencí. Tmavší stínovaná oblast nad křivkou označuje podmínky, kdy je pravděpodobné, že populace budou podle většiny konceptů považovány za samostatné druhy. Různé ochrany jednotka pojmy a infra-konkrétní taxonomické kategorie jsou zobrazeny ve vztahu k oběma osami genetické rozmanitosti (po Moritz, 2002).
to vede k otázce: proč ne jen pojmenovat ESUs jako poddruh? Poddruhy byly tradičně definovány a vnímány jak botanici a zoologové, jako fenotypově odlišné, allopatric sad populace, které mohou mezistupeň do sebe na zeměpisné hranice (Mayr, 1942; Grant, 1981). Jako takové potenciálně odrážejí adaptivní divergenci, aniž by nutně byly historicky izolovány. Označení poddruhů jsou široce používána v rostlinách, přičemž geografické a ekologické rozdíly jsou primárními prostředky, kterými se rozlišují (Hamilton a Reichard, 1992). Pokud poddruh projevovat lokální adaptace, s nebo bez probíhající genový tok, doporučujeme fenotypově definovanými poddruhy by měly i nadále být uznávána v zachování, a ne zaměnit s ESUs, alespoň ne Ježíši, jak je definován představují historicky izolované populace.
V stejným způsobem, jako je populační genomika poskytuje mnohem větší přesnost v identifikaci Ežíš využitím oba neutrální a adaptivní loci to také poskytuje prostředky pro využití velkého počtu neutrální loci, aby se více snadno vymezit Řízení Jednotek (MUs) jako demograficky nezávislé jednotky, vyznačující se tím i omezené tok genů (Funk et al., 2012). Zatímco většina z našich zaměření je zde na ESU a obdobné jednotky, jako jsou DUs, který může být rozpoznán jako hlavní intra-specifické jednotky pro ochranu a byly přijaty pod ochranu některých jurisdikcích, MUs byly identifikovány také představuje demograficky nezávislé soubory populací (viz Palsbøll et al., 2007), které lze zvládnout udržet větší ESU (Moritz, 1994; Funk et al., 2012). Řídící jednotky mohou být důležité při sledování a řízení populací druhů a byly také označovány jako „populace“, zvláště důležité pro řízení rybolovu (Avise, 2004; Laikre et al ., 2005). Definice MUs Moritz (1994) jako „populací s významným divergence alel v jaderné nebo mitochondriální lokusy, bez ohledu na fylogenetické rozlišovací způsobilosti alely“ je obecně široce přijímaný, i když, jak zdůraznil Palsbøll et al. (2007), demografická konektivita je klíčovým faktorem, nikoli úrovní historického toku genů.
Vymezují ochranná jednotky jako důležité prvky vnitrodruhových rozmanitost má také významný dopad na genetickou záchranu malých inbrední populace prostřednictvím obnovení toku genů nebo prsou, a to je stále více jasné, že genetická záchrana musí být šířeji, pokud se zvýšila populace zánik má být odvrácena (Láska Stowell et al., 2017; Ralls et al., 2018). Moritz (1999) navrhl, že míchání MUs, ale ne esu, by mohlo být považováno za vhodnou strategii pro genetickou záchranu. Jak zdůraznil Frankham et al. (2012) smíchání taxonomicky uznávaných druhů by manažeři obecně nepovažovali za přijatelné, v takovém případě by taxonomické přesazení mohlo omezit příležitosti k záchraně malých inbredních populací. Jsme naznačují, že zatímco přísnější přístup k vymezení vnitrodruhových jednotek jako MUs může pomoci zmírnit tento problém, to je také jasné, že genomické odhad míry migrace a adaptivní divergence má potenciál poskytnout mnohem více robustní vedení v nejen definování hranice druhů a vnitrodruhové variability, ale v návrhu genetické záchranné strategie.
Phylogenomics a Zachování
Genomika v kombinaci s fylogenetické metody lze snadno vyřešit vztahy a evoluční vzdálenost mezi taxony, stejně jako vymezení druhů. Toho lze nejlépe dosáhnout pomocí metod druhového stromu, spíše než zřetězení, jako první účet pro heterogenitu genového stromu a zabránit nadměrné inflaci délek hrotu v důsledku polymorfismu předků (Edwards et al ., 2016; Ogilvie a kol., 2016). Zatímco masivní počet lokusů často není podstatný, včetně více lokusů často zlepšuje rozlišení obou vztahů a délek větví (např., 2016). Význam pro ochranu přírody je dvojí. Za prvé, fylogenetické hodnocení umožňuje zvážit odlišnost druhů nebo ESUs, nejen to, zda jsou popsány. Program EDGE (Isaac et al., 2007) obhajuje vážení druhů pro ochranu na základě toho, kolik jedinečné evoluční historie představují, kromě obvyklých kritérií IUCN. Za druhé, samotné délky větví mohou představovat hodnoty biologické rozmanitosti, nezávislé na taxonomii (Faith, 1992), která prokázala hodnotu při upřednostňování oblastí ochrany (Laity et al ., 2015; Rosauer a kol., 2016, 2018). V tomto smyslu, přijetí fylogenetický metriky pomocí takových opatření, jako fylogenetická diverzita a fylogenetické endemismus, aby bylo možné upozornit na oblasti vysoké evoluční rozmanitost, která může být zaměřena na ochranu, a být nejvyšší prioritou v každém procesu plánování pro lepší zachování biologické rozmanitosti. Tento přístup může zvýšit ochranu genetické rozmanitosti a evolučních procesů pro různé skupiny organismů v celé řadě geografických měřítek (Laity et al., 2015) rozhodování o oblastech, které upřednostňují robustnější rozmary toho, jak jsou genomické důkazy přeloženy do rozpoznávání druhů nebo uvnitř specifických jednotek.
reálný svět-legislativní definice
politika a legislativní rámce pro ochranu biologické rozmanitosti sahají od mezinárodních úmluv a strategií, přes národní strategie a legislativu a státní nebo regionální legislativu a strategie v rámci zemí. Hlavních mezinárodních nástrojů na ochranu biologických druhů, patří Úmluva o Biologické Rozmanitosti realizován prostřednictvím Úmluvy smluvních Stran (COP), Úmluva o Světovém Dědictví, Globální Strategie v oblasti biologické Rozmanitosti, Montrealský Proces pro Zachování Lesů, Ramsarská Úmluva o Mokřadech, Úmluva o ochraně Stěhovavých Druhů, Úmluvě o Mezinárodním Obchodu Ohroženými Druhy (CITES), Mezinárodní Unie pro Ochranu Přírody (IUCN) v červeném seznamu ohrožených druhů, a na Rámcovou úmluvu organizace Spojených Národů o změně klimatu. Všechny tyto chartery ztělesňují hlavní, zachování druhů a genetické rozmanitosti jako základní cíl ochranu biologické rozmanitosti, s druhy jako jednotky široce vymezených pro ochranu. Definice druhů podle právních předpisů a mezinárodních směrnic jsou však velmi variabilní. Obsahují taxonomické rozpoznávání druhů a vnitrodruhových taxonů, ale může také zahrnovat genetické/evoluční pojmy, které jsou často zahrnuty pod právní definici „druhu“ (Tabulka 1).

Tabulka 1. Světové právní předpisy a směrnice, které definují „druhy“ pro zařazení a ochranu.
vzhledem k širokému spektru infra specifických úrovní, které lze chránit (Tabulka 1), by právní předpisy a související procesy zařazení zemí měly usnadnit zachování genetické rozmanitosti. Například definice „druhu“ , když vezmeme v úvahu legislativu po celém světě (Tabulka 1), může zahrnovat: druhy, poddruhy, odrůdy a geograficky a / nebo geneticky odlišné populace, zatímco podle jihoafrických právních předpisů mohou zahrnovat také kultivary, zeměpisné rasy, kmeny a hybridy. Tedy, ve většině právních předpisů po celém světě „druhů“ zahrnuje entity, které odrážejí různé infra-specifické limity, které jsou všechny součástí širší rozložení druhů a odrážejí různé míře populace-druhy kontinua. To se vyrovná s nevyhnutelnými obtížemi, kterým taxonomové, evoluční biologové a genetici ochrany čelí při hledání konzistentnějšího a jednotnějšího způsobu určení jednotek v tomto kontinuu.
někteří argumentují, že obavy týkající se politických důsledků druhů a infraspecifické nejistoty jsou na místě (Hej et al., 2003), ale stále jsme toho názoru, že ačkoli právní předpis může poskytnout mechanismus pro ochranu genetické diverzity v řadě úrovní, že důkazní břemeno je na biologové zajistit větší srozumitelnost a konzistenci na to, jak tyto úrovně by měly být odděleny od druhů prostřednictvím jednotlivých populací. V současné době jsou manažeři a zákonodárci ponecháni s matoucí řadou druhových konceptů a infraspecifických jednotek poskytovaných vědci při vymezení jednotek napříč kontinuem populačních druhů. Například, všech jurisdikcích zvážit druh uznán taxonomové, jako klíčových jednotek pro ochranu přírody (Tabulka 1), přesto je zde stále široké diskusi mezi taxonomové kolem druhy, pojmy, a to jak druhů jsou diagnostikovány v praxi se liší v rámci jednotlivých skupin. To vedlo k nedávné volání pro více regulovaný přístup k taxonomii (Garnett a Christidis, 2017), na které jiní namítali, že nařízení taxonomie není odpověď (Thomson et al., 2018). I když souhlasíme s některými body Thomson et al. (2018) navrhujeme, že je poněkud naivní navrhovat, aby se legislativa sama změnila, aby výslovně odkazovala na konkrétní koncept taxonu naznačený jménem. Tyto a předchozí podobné debaty zdůrazňují potřebu evoluční biologové a taxonomové, aby se ujala vedení v zajištění jasnosti (transparentnosti) v tom, jak se promítají genomické a další důkazy, o vymezení druhů nebo vnitrodruhových jednotek, a jak se tyto taxony, které se týkají příslušných právních předpisů pro zachování a ochranu.
Překlad pro Řízení
Jak bylo uvedeno výše, všech právních předpisů a směrnic rozpoznat úrovni druhu jako základní jednotky přírody, ale také zajistit uznávání různých infra-konkrétní jednotky. Mnozí mají také implicitní požadavek na formální pojmenování a taxonomický popis druhu, když za pro zachování priority a formální ochrany. Například, podle pokynů pro Červený seznam IUCN se nedoporučuje seznam nejmenovaných druhů a existuje požadavek, aby nový popis druhu byl zveřejněn do čtyř let od zařazení druhu na Červený seznam IUCN, nebo by byl odstraněn. To zdůrazňuje význam taxonomické pojmenování a popis subjektů, u obou druhů a infra-specifické úrovně, omezené taxonomické informace pro mnoho rostlinných a živočišných skupin (Mace, 2004; Oliver a Lee, 2010; Wilson, 2017), a že je třeba urychlit taxonomické úsilí, zejména v oblasti biologické rozmanitosti bohaté oblasti (Riedel et al., 2013; Wege et al., 2015; Wilson, 2017).
Taxonomicky jménem druhů a vnitrodruhových taxonomických kategorií, jako jsou poddruhy a odrůdy, obecně poskytuje základní jednotky pro formální zápis, zachování priorit a ochranu, ale v některých jurisdikcích jiné kategorie vnitrodruhových zachování jednotky jsou formálně uznána. Ty jsou obecně identifikovány pomocí kombinovaných genomických a fenotypových dat. Například zákon USA o ohrožených druzích uznává odlišné populační segmenty (DPSs), které jsou široce založeny na esu (Waples, 1991), zatímco Kanadský zákon o ohrožených druzích uznává DUs. Funkční výklad pojmu „zřetelný populace“ v rámci Kanadské Druhů ohrožených Zákona, DUs cílem identifikovat diskrétní sady populací, které představují důležité komponenty evolučního dědictví druhu a, které jsou nepravděpodobné, že by nahradit přirozený rozptyl (Mee et al., 2015).
DUs i DPSs se koncepčně překrývají s esu (Waples et al., 2013) a byly nejvíce aplikovány na severoamerické ryby, včetně lososovitých severozápadních USA (Waples, 1995) a kanadských koregonidů (Mee et al., 2015) je zajímavé, že DPSs v USA se vztahují pouze na obratlovce, zatímco v Kanadě může být DUs aplikován na jakýkoli organismus, který může být uveden podle jeho zákona (viz Waples et al ., 2013), s dusem v současné době uvedeným pro členovce, měkkýše a rostliny. V Austrálii, Kanadě, odlišné populace mohou být uvedeny pro každý organismus uznána v rámci Ochrany životního Prostředí a Zachování biologické Rozmanitosti z roku 1999 (EPBC Zákon), i když nejsou konkrétně uvedeny kategorie, jako DUSs a DPSs. Dosud však byly uznány pouze u obratlovců. V USA existuje 127 obratlovců DPSs uvedeny pro ochranu přírody (https://ecos.fws.gov/ecp/), v Kanadě 70 DUs byly uvedeny zahrnující všechny organismy (http://www.registrelep-sararegistry.gc.ca/species/schedules_e.cfm?id=1); a v Austrálii byly populace 7 druhů obratlovců pokrývajících savce, ryby a ptáky uznány za zvláštní stav ochrany podle zákona EPBC (http://www.environment.gov.au/cgi-bin/sprat/public/publicthreatenedlist.pl). Zatímco hodnocení těchto populací v USA a Kanadě obecně používá genetickou informaci (Waples et al., 2013), v Austrálii jsou obvykle založeny na geografických a teritoriálních identifikace, i když v některých z těchto případů byly podpořeny analýzou genetické diferenciace u savců (Firestone et al., 1999; Armstrong, 2006) a fish (Colgan a Paxton, 1997; Ahonen et al., 2009).
výpis infra-zvláštní ochrany jednotky pro rostliny a bezobratlé v Kanadě (DUs), ale ne v USA nebo v Austrálii ukazuje další problém není specificky týkající se vymezení ochranných jednotek, ale obecněji k významným nesrovnalostem v oblasti zachování biologické rozmanitosti priorit; kultovní obratlovců, jako jsou ptáci, savci a ryby jsou zaměření v mnoha jurisdikcích, zatímco ostatní organismy nejsou. Tento preferenční seznam populací určitých organismů odráží rozsah problémů, které jsou méně co do činění s vědou, které vymezují ochranu jednotek a více co do činění s výpis z populace na základě povědomí veřejnosti, politické účelnosti a charismatický charakter organismu, které mají být uvedeny (viz Waples et al., 2013).
Zatímco v Austrálii některých populací známých savců, ptáků a ryb jsou chráněny pod EPBC Zákon, tam je nyní důležité informace, na základě molekulárních studií u plazů, bezobratlých a rostlin, kde populace může být uznána jako ESUs nebo MUs, a mnozí z nich jsou z hlediska ochrany význam vzhledem k jejich úzké zeměpisné oblasti, malá velikost populace, míra poklesu, a náchylnost k ohrožující procesy. Například, stávkující příklady z vysoce strukturované populace a hluboce odlišných linií v rámci druhu, které by se snadno vešly požadavek na ESU lze nalézt v řadě nízké rozptýlení obratlovců z tropů (Obrázek 2) a jinde. Podobně v Australské bezobratlých, jako jsou různonožci a mnohonožky, významné genetické struktury, a phylogeographic vzory podporují vymezení diskrétní populace klastry jako ESUs, které jsou geograficky nesourodé, s úzkými zeměpisné oblasti, omezené na konkrétní stanoviště a zastoupena pouze několika populací (Finston et al., 2007; Murphy et al., 2013; Nistelberger a kol., 2014a). Často jsou tyto endemické krátkého dosahu taxony ochrany (Harvey et al ., 2011) a jsou společně umístěny v hotspotech endemismu (např., 2017), které jsou samy o sobě vysokými prioritami pro ochranu více taxonů (Rosauer et al ., 2018), jako je oblast Kimberley a Stirling Range v jihozápadní západní Austrálii.
V Australské rostliny tam jsou také značné množství populace genetické a phylogeographic studie identifikace odlišných skupin obyvatelstva v rámci řady druhů z různých rodů odráží disjunktní a historicky izolované populace systémů, geologických a půdních složitosti, a zabírá kontrastní stanoviště z hlediska vegetace a klimatu (viz Broadhurst et al., 2017). Geneticky odlišné populace jsou zvláště patrné u druhů z jihozápadní západní Austrálie (Coates, 2000; Byrne et al., 2014) s řadou studie zdůrazňující významnou genetickou strukturu a velmi odlišných linií v řadě rostlinných rodů a rodin, včetně: Akát (Mimosaceae; Coates, 1988), Atriplex (Amaranthaceae; Shepherd et al., 2015), Banksia (Proteaceae; Coates et al., 2015), Lambertia (Proteaceae; Byrne et al., 2002), Hakea (Proteaceae; Sampson et al., 2014), Eremophila (Scrophulariaceae; Llorens et al., 2015), Pultanaea (Fabaceae; Millar a Byrne, 2013) a Calothamnus (Myrtaceae; Nistelberger et al., 2014b). V mnoha případech jsou tyto populace uznány při plánování ochrany státními vládními agenturami a zohledněny při posuzování dopadů na životní prostředí (viz Shepherd et al., 2015) i když nejsou uvedeny pro ochranu podle právních předpisů. Uznáváme však, že formální uznání těchto jednotek by z dlouhodobého hlediska zlepšilo jejich zachování.
tyto australské příklady zdůrazňují rostoucí potřebu rozpoznat tuto vrstvu genetické rozmanitosti pod úrovní druhů. Úkolem je zajistit, aby vysoce strukturované populace a hluboce odlišné linie, které jsou předmětem ochrany, byly chráněny tváří v tvář konkurenčním požadavkům na ochranu. Kromě ikonických druhů je tato výzva patrná i celosvětově, s výjimkou Kanady. Neschopnost chránit jasně vymezena ochranná jednotky bude pravděpodobně zvyšovat riziko záhadné vymírání a ztráty významné genetické rozmanitosti, což v mnoha případech může zahrnovat rody a historicky izolované populace, které přetrvávají v rámci druhu po miliony let (Moritz, 2002).
Závěry a Doporučení
seznam druhů a různé infra-zvláštní kategorie pod ohrožených druhů právních předpisů nebo podle mezinárodní organizace jako IUCN, označuje závazek k ochraně genetické rozmanitosti napříč druhy, populace kontinua. Přesto současné přístupy k ochraně biologické rozmanitosti do značné míry zaměřit se na druhy konec tohoto kontinua s menší pozornost věnována genetické rozmanitosti a infra-specifické rozdíly. Tento více omezené zaměření na vnitrodruhové úrovni genetické variability naznačuje, že při zachování manažeři stále více uvědomují hodnotu explicitně zvažuje genetické informace v řízení akcí a priorit, často jim chybí žalovatelné informace z rozmanitosti vědců (Waples et al., 2008; Laikre a kol., 2010; Cook a Sgrò, 2017; Taylor a kol., 2017). Tato mezera mezi vědou a konzervační praxí brání účinnějšímu globálnímu přístupu k zachování a řízení genetické rozmanitosti.
navrhujeme, že nedostatek jasnosti vědců o biologické rozmanitosti nad jednotkami ochrany je do značné míry založen na nesrovnalostech na dvou úrovních. První se týká konceptů více druhů a různých diagnostických metod používaných k vymezení druhů, které vytvářejí nekonzistenci napříč taxonomickými skupinami. Druhý se týká nedostatek konzistence a dohody taxonomové, evoluční biologové, a zachování biologů o tom, jak zacházet infra-specifické taxony a další subjekty, jako jsou Ježíš, DUs, a MUs.
Když vezmeme v úvahu, že právní předpisy jsou často již dostatečně flexibilní, pokud jde o to, jak taxonů („druhů“) jsou definovány, můžeme tvrdit, že je třeba systematicky řešit populace druhů kontinua, a jasněji definovat jednotky pro zachování větší uznání pojmy jako ESU je a jak se tyto týkají příslušné právní předpisy. Zdůrazňujeme, že tato otázka bude bezpochyby kritičtější vzhledem k síle genomiky detekovat diferenciaci v postupně jemnějších měřítcích a její rostoucí využití k vymezení druhů a infraspecifických entit.
Z předchozí analýzy, nabízíme následující doporučení:
(i) Genomických dat by měly být interpretovány obezřetně při vymezování druhů, pomocí integrační taxonomický přístup, který obvykle zahrnuje důkazy, mimo genetické údaje.
(ii), Kde genetici určit více linií nebo odlišné populace v rámci druhu, a tyto nezaručují nadmořská výška na úrovni druhu, a nejsou taxonomicky poznal, že tam být silnější impuls a více jednotný přístup k identifikaci jasně zdůvodněno vnitrodruhových jednotek pro praktické posouzení zachování stavu nebo dopadů, a že tyto být výslovně uznány jako jednotky pro zachování zdrojů v souvislosti s právními předpisy dané jurisdikce.
(iii) zvážit podle IUCN, a to prostřednictvím jeho Zachování Genetika Specialista Skupiny a její regionální sub-skupin, vypracování pokynů pro rozpoznání a označování vnitrodruhových jednotek, které sladit s nejlepší vědecké praxi.
(iv) podrobnější pohled vznikající z evoluční genomika druhů a kontinuum od populací druhu by měly být sdělovány efektivněji tím, že vědci k zachování manažerů a politiků.
obecněji opakujeme doporučení učiněná nedávno v řadě recenzí (Frankham, 2010; Santamaría and Méndez, 2012; Hoban et al., 2013; Moritz a Potter, 2013; Laity a kol., 2015; Pierson a kol., 2016; Cook a Sgrò, 2017; Taylor a kol., 2017), které zdůrazňují nutnost vědci ke zlepšení komunikace a zvyšování povědomí s tvůrci politiky, vládních agentur, nevládních organizací a lékařů, že výsledky z molekulární systematika, molekulární taxonomie, fylogeografie a populační genetický výzkum může poskytnout důležité informace o zachování genetické rozmanitosti a řízení evolučních procesů.
autorské příspěvky
DC, MB a CM přispěly k přípravě a revizi rukopisu.
Prohlášení o střetu zájmů
autoři prohlašují, že výzkum byl proveden bez jakýchkoli obchodních nebo finančních vztahů, které by mohly být vykládány jako potenciální střet zájmů.
poděkování
Tento projekt byl podpořen australskou Radou pro výzkum (ARC Linkage Grant No. LP120200063), Australská národní univerzita a západní australské Oddělení biologické rozmanitosti, ochrany a atrakcí.
Afonso Silva, a. C., Braggová, J. G., Potter, S., Fernandes, C., Coelho, M. M., a Moritz, C. (2017a). Tropický specialista versus klimatolog: diverzifikace a demografická historie sesterských druhů carlia skinks ze severozápadní Austrálie. Molo. Ecol. 26:4045–4058. doi: 10.1111 / mec.14185
CrossRef Plný Text | Google Scholar
Afonso Silva, a. C., Santos, N., Ogilvie, H. a., a Moritz, C. (2017b). Validace a popis dvou nových severozápadních australských duhových skinků s multidruhovými koalescent metodami a morfologií. PeerJ 5: e3724. doi: 10.7717/peerj.3724
PubMed Abstract / CrossRef Full Text / Google Scholar
Ahonen, h., Harcourt, R. G., and Stow, a. J. (2009). Jaderná a mitochondriální DNA odhaluje izolaci ohrožených populací žraloků šedých (Carcharias taurus). Molo. Ecol. 18, 4409–4421. doi: 10.1111 / j. 1365-294X. 2009. 04377.x
PubMed Abstract / CrossRef Full Text / Google Scholar
Armstrong, k. n. (2006). Fylogeografická struktura u Rhinonicteris aurantia (Chiroptera: Hipposideridae): důsledky pro ochranu. Acta Chiropterologica 8, 63-81. doi: 10.3161/1733-5329(2006)82.0.CO
CrossRef Plný Text | Google Scholar
Arnold, M. L., a Kunte, K. (2017). Adaptivní genetická výměna: zamotaná historie příměsí a evoluční inovace. Trendy Ecol. Evol. 32, 601–611. doi: 10.1016 / j. strom.2017.05.007
PubMed Abstract / CrossRef Full Text / Google Scholar
Avise, J. C. (2004). Molekulární markery, přírodní historie a evoluce. 2.Edn. Sunderland, MA; Oxford: Oxford University Press.
Google Scholar
Avise, J. C., Arnold, J., Ball, R. M., Bermingham, e., Lamb, T., Neigel, J. E., et al. (1987). Intraspecifická fylogeografie: mitochondriální DNA most mezi populační genetikou a systematikou. Anna. Rev. Ecol. Sys. 18, 489–522. doi: 10.1146/annurev.es.18.110187.002421
CrossRef Plný Text | Google Scholar
Bickford, D., Lohman, D. J., Sodhi, N, Ng, P. K., Meier, R., Mrkat, K., et al. (2007). Kryptické druhy jako okno rozmanitosti a ochrany. Trendy Ecol. Evol. 22, 148–155. doi: 10.1016 / j. strom.2006.11.004
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Blom, M. P. K., Bragg, J. G., Potter, S., a Moritz, C. (2016). Účtování nejistoty v odhadu genového stromu: shrnutí-koalescentní odvození stromu druhů v náročném záření australských ještěrek. Syst. Biol. 66, 352–366. doi: 10.1093/sysbio/syw089
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Broadhurst, L., Plemeno, M., Lowe, V., Bragg, J., Catullo, R., Coates, D. J., et al. (2017). Genetická rozmanitost a struktura australské flóry. Různý. Distrib. 23, 41–52. doi: 10.1111 / ddi.12505
CrossRef Full Text / Google Scholar
Byrne, m., Coates, D. J., Forest, F., Hopper, S. D., Krauss, S. L., Sniderman, J. M. K., et al. (2014). „Rozmanitá flora – druhy a genetické vztahy,“ v Životě Rostlin na Sandplains v Jihozápadní Austrálii, Globální biologické Rozmanitosti Hotspot, redakce H. Lambers (Crawley, WA: University of Western Australia Publikování), 81-99.
PubMed Abstract
Byrne, m., Macdonald, B., and Coates, D. J. (2002). Fylogeografické vzory a vysoká rozmanitost cpDNA v rámci Acacia acuminata (Leguminosae: Mimosoideae). komplex v Západní Austrálii. J.Evol. Biol. 15, 576–587. doi: 10.1046 / j. 1420-9101. 2002. 00429.x
CrossRef Plný Text | Google Scholar
Carstens, B. C., Pelletier, T. a., Reid, N. M., a Satler, J. D. (2013). Jak selhat při vymezení druhů. Molo. Ecol. 22, 4369–4383. doi: 10.1111 / mec.12413
PubMed Abstract | CrossRef Full Text | Google Scholar
Coates, D.J. (1988). Genetická rozmanitost a struktura populačního genu ve vzácném Chitteringu trávy, akácie anomala Court. Aust. J. Bot. 36:273–286. doi: 10.1071/BT9880273
CrossRef Full Text | Google Scholar
Coates, D.J. (2000). Definování ochranářských jednotek v bohaté a roztříštěné flóře: důsledky pro řízení genetických zdrojů a evolučních procesů v rostlinách jihozápadní Austrálie. Aust. J. Bot. 48, 329–339. doi: 10.1071/BT99018
CrossRef Plný Text | Google Scholar
Coates, D. J., McArthur, S. L., Byrne, M. (2015). Významná ztráta genetické rozmanitosti po vyhynutí populace způsobené patogenem u vzácné endemické Banksia brownii (Proteaceae). Biol. Konzervativec. 192, 353–360. doi: 10.1016 / j. biocon.2015.10.013
CrossRef Full Text | Google Scholar
Colgan, D.J., and Paxton, J. R. (1997). Biochemická genetika a rozpoznání západní populace gemfish obecné, Rexea solandri (Scombroidea: Gempylidae)., v Austrálii. Březen. Freshw. Rez.48, 103-118. doi: 10.1071/MF96048
CrossRef Plný Text | Google Scholar
Úmluva o Biologické Rozmanitosti (2007). K dispozici online na adrese: https://www.cbd.int/doc/meetings/cop-bureau/cop-bur-2007/cop-bur-2007-10-14-en.pdf
Cook, C. N., and Sgrò, C. M. (2017). Sladění vědy a politiky s cílem dosáhnout evolučně osvícené ochrany přírody. Konzervativec. Biol. 31, 501–512. doi: 10.1111/cobi.12863
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Cook, L. G., Edwards, R. D., Ostrý, M. D., a Hardy, N. B. (2010). Potřebujete morfologii vždy vyžadovanou pro nové popisy druhů? Invertovat. Sys. 24, 322–332. doi: 10.1071/IS10011
CrossRef Plný Text | Google Scholar
Cracraft, J. (1983). Druhové pojmy a speciační analýza. Curre. Ornithol. 1, 159–187. doi: 10.1007/978-1-4615-6781-3_6
CrossRef Plný Text | Google Scholar
Crandall, K. a., Bininda-Emonds, O. R., muškátový květ, G. M., a Wayne, R. K. (2000). Vzhledem k evolučním procesům v biologii ochrany. Trendy Ecol. Evol. 15, 290–295. doi: 10.1016/S0169-5347(00)01876-0
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Frézy, a. D., a Payseur, B. a. (2013). Genomické podpisy výběru na odkazovaných místech: Sjednocení rozdílů mezi druhy. Adresa. Reverend Genet. 14, 262–274. doi: 10.1038/nrg3425
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Dayrat, B. (2005). Směrem k integrační taxonomii. Biol. J. Linn. SOC. 85, 407–415. doi: 10.1111 / j. 1095-8312. 2005. 00503.x5
CrossRef Full Text / Google Scholar
de Queiroz, k. (1998). „Generální linie konceptu druhů, druhů kritérií a procesu speciace“, ve Formách: Druhy a Speciace, D. J. Howard,r. S. H. Berlocher (New York, NY: Oxford University Press), 57-75.
Google Scholar
de Queiroz, k. (2007). Druhové pojmy a vymezení druhů. Syst. Biol. 56, 879–886. doi: 10.1080/10635150701701083
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Degnan, J. H., a Rosenberg, N. a. (2009). Nesoulad genového stromu, fylogenetická inference a vícedruhové koalescence. Trendy Ecol. Evol. 24, 332–340. doi: 10.1016 / j. strom.2009.01.009
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Devey, D. S., Bateman, R. M., Fay, M. F., a Hawkins, J. a. (2008). Přátelé nebo příbuzní? Fylogenetika a vymezení druhů v kontroverzním evropském rodu orchidejí Ophrys. Anna. Bota. 101, 385–402. doi: 10.1093/aob/mcm299
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Dizon, a. E., Lockyer, C, Perrin, W. F., Demaster, D. P., a Sisson, J. (1992). Přehodnocení konceptu akcií-fylogeografický přístup. Konzervativec. Biol. 6, 24–36. doi: 10.1046 / j. 1523-1739. 1992. 610024.x
CrossRef Plný Text | Google Scholar
Doughty, P., Bourke, G. Tedeschi, L., Pratt, R., Oliver, P., Palmer, R., et al. (2018). Vymezení druhů v Gehyra nana (Squamata: Gekkonidae). komplex: kryptický a divergentní morfologický vývoj v australských monzunových tropech s popisem čtyř nových druhů. Zootaxa, 4403: 201-244. doi: 10.11646 / zootaxa.4403.2.1
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Dynesius, M., a Jansson, R. (2014). Perzistence linií uvnitř druhů: zanedbaná kontrola míry speciace. Evoluce 68, 923-934. doi: 10.1111 / evo.12316
PubMed Abstract / CrossRef Full Text / Google Scholar
Edwards, s. v.,Potter, s., Schmitt, C. J., Bragg, J. G., and Moritz, C. (2016). Retikulace, divergence a fylogeografie–fylogenetické kontinuumc. Proc. Natle. Acad. Věda. U. S. A. 113, 8025-8032. doi: 10.1073 / pnas.1601066113
CrossRef Full Text | Google Scholar
Faith, D. P. (1992). Hodnocení zachování a fylogenetická rozmanitost. Biol. Nevýhoda. 61, 1–10. doi: 10.1016/0006-3207(92)91201-3
CrossRef Plný Text | Google Scholar
Finston, T. L., Johnson, M. S., Humphreys, W. F., Eberhard, S. M., a Halse, S. a. (2007). Kryptická speciace ve dvou rozšířených podzemních rodech amphipod odráží historické drenážní vzorce ve starověké krajině. Molo. Ecol. 16, 355–365. doi: 10.1111 / j. 1365-294X. 2006. 03123.x
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Firestone, K. B., Elphinstone, M. S., Sherwin, W. B., a Houlden, B. a. (1999). Fylogeografická populační struktura tygra quolls Dasyurus maculatus (Dasyuridae: Marsupalia), ohroženého masožravého vačnatce. Molo. Ecol. 8, 1613–1625. doi: 10.1046 / j. 1365-294x. 1999. 00745.x
CrossRef Full Text / Google Scholar
Fišer, C., Robinson, C. T., and Malard, F. (2018). Kryptické druhy jako okno do posunu paradigmatu druhového konceptu. Molo. Ecol. 27, 613–635. doi: 10.1111 / mec.14486
PubMed Abstract / CrossRef Full Text / Google Scholar
Frankham, R. (2010). Výzvy a příležitosti genetických přístupů k biologické ochraně. Biol. Konzervativec. 143,1919–1927. doi: 10.1016 / j. biocon.2010.05.011
CrossRef Plný Text | Google Scholar
Frankham, R., Ballou, J. D., Dudash, M. R., Eldridge, M. D., Fenster, C. B., Lacy, R. C., et al. (2012). Důsledky různých druhových konceptů pro zachování biologické rozmanitosti. Biol. Konzervativec. 153, 25–31. doi: 10.1016 / j. biocon.2012.04.034
CrossRef Plný Text | Google Scholar
Fraser, D. J., a Bernatchez, L. (2001). Adaptivní evoluční ochrana: směrem k jednotnému konceptu definování ochranářských jednotek. Molo. Ecol. 10, 2741–2752. doi: 10.1046 / j. 1365-294X. 2001.t01-1-01411.x
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Funk, W. C., Mckay, J. K., Hohenlohe, P. a., and Allendorf, F.W. (2012). Využití genomiky pro vymezení ochranářských jednotek. Trendy Ecol. Evol. 27, 489–496. doi: 10.1016 / j. strom.2012.05.012
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Garnett, S. T., a Christidis, L. (2017). Taxonomie anarchie brání zachování. Příroda 546, 25-27. doi: 10.1038/546025a
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Gippoliti, S., Cotterill, F. P. D., Zinner, D., a Groves, C. P. (2018). Dopady taxonomické setrvačnosti na zachování rozmanitosti afrických kopytníků: Přehled. Biol. Rev. 93, 115-130. doi: 10.1111 / brv.12335
PubMed Abstract | CrossRef Full Text | Google Scholar
Grant, V. (1981). Speciace Rostlin. New York, NY: Columbia University Press.
PubMed Abstract / Google Scholar
Hamilton, CW, and Reichard, S. H. (1992). Současná praxe v používání poddruhů, odrůd a forem v klasifikaci divokých rostlin. Taxon 41, 485-498. doi: 10.2307 / 1222819
CrossRef Full Text / Google Scholar
Harvey, MS, Rix, M. G., Framenau, v. w., Hamilton, z. R., Johnson, M. S., Teale, R. J., et al. (2011). Ochrana nevinných: studium endemických taxonů krátkého dosahu zvyšuje výsledky ochrany. Invertovat. Syst. 25, 1–10. doi: 10.1071/IS11011
CrossRef Plný Text | Google Scholar
Hej, J., Waples, R. S., Arnold, M. L., Butlin, R. K., a. Harrison, R. G. (2003). Porozumění a konfrontace druhové nejistoty v biologii a ochraně. Trendy Ecol. Evol. 18, 597–603. doi: 10.1016 / j. strom.2003.08.014
CrossRef Plný Text | Google Scholar
Hoban, S. M., Hauffe, H. C., Perez-Espona, S., Arntzen, J. W., Bertorelle, G., Bryja, J., et al. (2013). Uvedení genetické rozmanitosti do popředí politiky a řízení ochrany přírody. Konzervativec. Genete. Resour. 5, 593–598. doi: 10.1007/s12686-013-9859-y
CrossRef Plný Text | Google Scholar
Isaac, N. J., Palička, J., a muškátový květ, G. M. (2004). Taxonomická inflace: její vliv na makroekologii a zachování. Trendy Ecol. Evol. 19, 464–469. doi: 10.1016 / j. strom.2004.06.004
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Isaac, N. J., Turvey, S. T., Collen, B., Waterman, C., a Fakulty, J. E. (2007). Savci na okraji: priority ochrany založené na hrozbě a fylogenezi. PLoS ONE 2: e296. doi: 10.1371 / deník.pone.0000296
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Jones, M. R., Dobrý, J. M. (2016). Cílené zachycení v evoluční a ekologické genomice. Molo. Ecol. 25, 185–202. doi: 10.1111 / mec.13304
PubMed Abstract / CrossRef Full Text / Google Scholar
Jorger, K. M., and Schrödl, m. (2013). Jak popsat záhadný druh? Praktické výzvy molekulární taxonomie. Před. Zool. 10:59. doi: 10.1186/1742-9994-10-59
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Laikre, L., Allendorf, F., Aroner, L. C., Baker, S., Gregovich, D. P., Hansen, M. M., et al. (2010). Zanedbání genetické rozmanitosti při provádění Úmluvy o biologické rozmanitosti. Konzervativec. Biol. 24, 86–88. doi: 10.1111 / j. 1523-1739. 2009. 01425.x
PubMed Abstract / CrossRef Full Text / Google Scholar
Laikre, L., Palm, s., and Ryman, N. (2005). Genetická populační struktura ryb: důsledky pro správu pobřežních zón. Ambio 34,111-119. doi: 10.1579/0044-7447-34.2.111
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Laici, T., Laffan, S. W., González-Orozco, C. E., Víra, D. P., Rosauer, D. F., Byrne, M., et al. (2015). Fylodiverzita k informování o ochraně přírody: Australský příklad. Věda. Celkové Prostředí. 534, 131–143. doi: 10.1016 / j. scitotenv.2015.04.113
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Leache, a. D., a Oaks, J. R. (2017). Užitečnost jednonukleotidového polymorfismu (SNP). data ve fylogenetice. Anna. Rev. Ecol. Syst. 48, 69–84. doi: 10.1146/annurev-ecolsys-110316-022645
CrossRef Full Text | Google Scholar
Llorens, T. M., Macdonald, B., McArthur, S., Coates, D. J., and Byrne, M. (2015). Disjunct, highly divergent genetic lineages within two rare Eremophila (Scrophulariaceae: Myoporeae). species in a biodiversity hotspot: implications for taxonomy and conservation. Biol. J. Linn. Soc. 177, 96–111. doi: 10.1111/boj.12228
CrossRef Full Text | Google Scholar
Love Stowell, S. M., Pinzone, C. A., and Martin, A. P. (2017). Překonávání překážek aktivních zásahů do genetické rozmanitosti. Biodivers. Konzervativec. 26, 1753–1765. doi: 10.1007/s10531-017-1330-z
CrossRef Plný Text | Google Scholar
Mace, G. M. (2004). Role taxonomie v ochraně druhů. Philos. Trans. R.Soc. B: Biol. Věda. 359, 711–719. doi: 10.1098 / rstb.2003.1454
PubMed Abstract / CrossRef Full Text / Google Scholar
Mallet, J. (2007). Hybridní speciace. Příroda 446, 279-283. doi: 10.1038 / nature05706
PubMed Abstract / CrossRef Full Text / Google Scholar
Mayr, e. (1942). Systematika a Původ druhů. New York, NY: Columbia University Press.
Google Scholar
Mee, J. a., Bernatchez, L., Reist, J. D., Rogers, S. M., a Taylor, E. B. (2015). Identifikace identifikovatelných jednotek pro stanovení priorit intraspecifické ochrany: hierarchický přístup aplikovaný na komplex druhů bílých ryb (Coregonus spp.). Evol. Appl. 8, 423–441. doi: 10.1111 / eva.12247
PubMed Abstract | CrossRef Full Text | Google Scholar
Millar, M. A., and Byrne, M. (2013). Cryptic divergent lineages of Pultenaea pauciflora M.B. Scott (Fabaceae: Mirbelieae) exhibit different evolutionary history. Biol. J. Linn. Soc. 108, 871–881. doi: 10.1111/bij.12002
CrossRef Full Text | Google Scholar
Moritz, C. (1994). Defining evolutionarily significant units for conservation. Trends Ecol. Evol. 9, 373–375. doi: 10.1016/0169-5347(94)90057-4
CrossRef Full Text | PubMed Abstract
Moritz, C. (1999). Konzervační jednotky a translokace: strategie pro zachování evolučních procesů. Hereditas 130, 217-228. doi: 10.1111 / j. 1601-5223. 1999. 00217.x
CrossRef Full Text / Google Scholar
Moritz, C. (2002). Strategie na ochranu biologické rozmanitosti a evolučních procesů, které ji udržují. Syst. Biol. 51, 238–254. doi: 10.1080/10635150252899752
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Moritz, C., a Potter, S. (2013). Význam evoluční perspektivy při plánování politiky ochrany. Molo. Ecol. 22, 5969–5971. doi: 10.1111 / mec.12565
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Moritz, C., Pratt, R. C., Banky, a. S., Bourke, G., Bragg, J. G., Doughty, P., et al. (2018). Kryptická rozmanitost linie, divergence velikosti těla a sympatry v druhovém komplexu australských ještěrů (Gehyra). Evoluce 27, 54-66. doi: 10.1111 / evo.13380
CrossRef Plný Text | Google Scholar
Morrison, W. R., Lohr, J. L., Duchen, P., Wilches, R., Trujillo, D., Mair, M., et al. (2009). Dopad taxonomické změny na zachování: Zabíjí, může zachránit, nebo je to jen irelevantní? Biol. Nevýhoda. 142, 3201–3206. doi: 10.1016 / j. biocon.2009.07.019
CrossRef Full Text / Google Scholar
Murphy, N. P., Adams, m., Guzik, M. T., and Austin, a.D. (2013). Mimořádný mikroendemismus v australských pouštních jarních amfipodech. Molo. Fylogenet. Evol. 66, 645–653. doi: 10.1016 / j. ympev.2012.10.013
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Murphy, N. P., Král, R. a., a Delean, S. (2015). Druhy, ESUs nebo populace? Vymezení a popis morfologicky kryptické rozmanitosti v australských pouštních jarních amfipodech. Invertovat. Syst. 29, 457–467. doi: 10.1071/IS14036
CrossRef Plný Text | Google Scholar
Nátěr, A., Avalon-Greminger, M. P., Nurcahyo, A., Nowak, M. G., de Ruční, M., Desai, T., et al. (2017). Morfometrické, behaviorální a genomické důkazy pro nový druh orangutana. Curre. Cell Biol. 27, 3487–3498. doi: 10.1016 / j. mládě.2017.09.047
CrossRef Plný Text | Google Scholar
Nicotra, a. B., Chong, C., Braggová, J. G., Ong, C. R., Aitken, s. N. C., Chuah, A., a kol. (2016). Populace a fylogenomický rozklad pomocí genotypizace sekvenováním v australském Pelargoniu. Molo. Ecol. 25, 2000–2014. doi: 10.1111 / mec.13584
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Nistelberger, H., Byrne, M., Coates, D., Roberts, J. D. (2014a). Silný phylogeographic struktury v stonožka označuje Pleistocénu vicariance mezi populacemi na páskované železné formace v semi-aridní Austrálie. PLoS ONE 9: e0093038. doi: 10.1371 / deník.pone.0093038
PubMed Abstract | CrossRef Full Text | Google Scholar
Nistelberger, h., Gibson, N., Macdonald, B., Tapper, S.-L., and Byrne, m. (2014b). Fylogeografický důkaz pro dvě mesic refugia v hotspotu biologické rozmanitosti. Dědičnost 113, 454-463. doi: 10.1038/hdy.2014.46
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Ogilvie, H. a., Heled, J., Xie, D., a Drummond, a. J. (2016). Výpočetní výkon a statistická přesnost beast a srovnání s jinými metodami. Syst. Biol. 65, 381–396. doi: 10.1093/sysbio/syv118
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Oliver, P. M., Keogh, J. S., a Moritx, C. (2015). Nové přístupy k katalogizaci a porozumění evoluční rozmanitosti: perspektiva australské herpetologie. Aust. J. Zool. 62, 417–430. doi: 10.1071/ZO14091
CrossRef Plný Text | Google Scholar
Oliver, P. M., Laver, R. J., Demello Martins, F., Pratt, R. C., Hunjan, a. S., a Moritz, C. (2017). Nový hotspot endemismu obratlovců a evoluční refugium v tropické Austrálii. Potápěč. Distrib. 23, 53–66. doi: 10.1111 / ddi.12506
CrossRef Plný Text | Google Scholar
Oliver, P. M., Lee, M. S. Y. (2010). Botanické a zoologické kódy brání výzkumu biologické rozmanitosti tím, že odrazují od zveřejnění nejmenovaných nových druhů. Taxon 59, 1201-205. doi: 10.2307/20773990
CrossRef Plný Text | Google Scholar
Padial, J., Miralles, A., De la Riva, I., a Vences, M. (2010). Integrační budoucnost taxonomie. Před. Zool. 7:16. doi: 10.1186/1742-9994-7-16
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Palsbøll, P. J., Bérubé, M., a Allendorf, F. W. (2007). Identifikace řídících jednotek pomocí populačních genetických dat. Trendy Ecol. Evol. 22, 11–16. doi: 10.1016 / j. strom.2006.09.003
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Patton, J. L. a Smith, M. F. (1994). Parafyly, polyfyly a povaha druhových hranic u kapes (Rod Thomomys). Syst. Biol. 43, 11–26. doi: 10.1093 / sysbio / 43.1.11.
CrossRef Plný Text | Google Scholar
Pierson, J. C., Coates, D. J., Oostermeijer, J. G. B., Beissinger, S. R., Bragg, J. G., Sunnucks, P., et al. (2016). Genetické faktory v plánech obnovy ohrožených druhů na třech kontinentech. Před. Ecol. Environ. 14, 433–440. doi: 10.1002 / poplatek.1323
CrossRef Plný Text | Google Scholar
Potter, S., Bragg, J. G., Petr, B. M., Bi, K., a Moritz, C. (2016). Fylogenomika na špičkách: odvozování linií a jejich demografické historie u tropické ještěrky, Carlia amax. Molo. Ecol. 25, 1367–1380. doi: 10.1111/Seč.13546
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Potter, S., Xue, a. T., Bragg, J. G., Rosauer, D. F., Roycroft, E. J., a Moritz, C. (2018). Pleistocénní klimatické změny vedou k diverzifikaci tropické savany. Molo. Ecol. 27, 520–532. doi: 10.1111 / mec.14441/plný
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Rabosky, D. L., Hutchinson, M. N., Donnellan, S. C., Talaba, A. L., a Lovette, I. J. (2014). Fylogenetická demontáž hranic druhů v rozšířené skupině australských skinků (Scincidae: Ctenotus). Molo. Fylogenet. Evol. 77, 71–82. doi: 10.1016 / j. ympev.2014.03.026
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Ralls, K., Ballou, J. D., Dudash, M. R., Eldridge, M. D. B., Fenster, C. B., Lacy, R. C., et al. (2018). Výzva k posunu paradigmatu v genetickém řízení roztříštěných populací. Konzervativec. Lette. 11, 1–6. doi: 10.1111 / konl.124126
CrossRef Full Text / Google Scholar
Rannala, B. (2015). Umění a věda o vymezení druhů. Curre. Zool. 61, 846–853. doi: 10.1093/czoolo/61.5.846
CrossRef Plný Text | Google Scholar
Riedel, A., Sagata, K., Suhardjono, Y. R., Tänzler, R., a Balke, M. (2013). Integrační taxonomie na rychlé cestě-směrem k větší udržitelnosti ve výzkumu biologické rozmanitosti. Před. Zool. 10:15. doi: 10.1186/1742-9994-10-15
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Rieseberg, L. H., a Willis, J. H. (2007). Speciace rostlin. Věda 317, 910-914. doi: 10.1126 / věda.1137729
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Rosauer, D. F., Blom, M. P. K., Bourke, G., Catalano, a. S., Donnellan, S., Mulder, E., et al. (2016). Fylogeografie, hotspoty a priority ochrany: příklad z horního konce Austrálie. Biol. Nevýhoda. 204, 83–93. doi: 10.1016 / j. biocon.2016.002
CrossRef Plný Text | Google Scholar
Rosauer, D. F., Byrne, M., Blom, M. P. K., Coates, D. J., Donnellan, S., Doughty, P., et al. (2018). Plánování ochrany v reálném světě pro evoluční rozmanitost v Kimberley, Austrálie, obchází nejistou konzervaci taxonomie. Lette. 11: e12438. doi: 10.1111 / konl.12438
CrossRef Plný Text | Google Scholar
Rosenblum, E. B., Sarver, B. a. J., Brown, J. W., Des Roches, S., Hardwick, K. M., Umožňují, T. D., et al. (2012). Zlatovláska splňuje Santa Rosalia: pomíjivý speciační model vysvětluje vzorce diverzifikace napříč časovými měřítky. Evol. Biol. 39, 255–261. doi: 10.1007/s11692-012-9171-x
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Roux, C., Fraïsse, C., Romiguier, J., Anciaux, Y., Galtier, N., a Bierne, N. (2016). Vrhá světlo na šedou zónu speciace podél kontinua genomové divergence. PLoS Biol. 14: e2000234. doi: 10.1371 / deník.pbio.2000234
PubMed Abstract / CrossRef Full Text / Google Scholar
Ryder, o.a. (1986). Ochrana druhů a systematika-dilema poddruhů. Trendy Ecol. Evol. 1, 9–10. doi: 10.1016/0169-5347(86)90059-5
CrossRef Plný Text | Google Scholar
Sampson, J. F., Byrne, M., Yates, C. J., Gibson, N., Thavornkanlapachai, R., Stankowski, a. S., et al. (2014). Současná genová imigrace zprostředkovaná pylem odráží historickou izolaci vzácného keře v roztříštěné krajině. Dědičnost 112, 172-181. doi: 10.1038/hdy.2013.89
PubMed Abstract / CrossRef Full Text / Google Scholar
Santamaría, L., and Méndez, P. F. (2012). Vývoj politiky biologické rozmanitosti: současné mezery a budoucí potřeby. Evol. Appl. 5, 202–218. doi: 10.1111 / j. 1752-4571. 2011. 00229.x
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Schlick-Steiner, B. C., Steiner, F. M., Seifert, B., Stauffer, C., Christiane, E., a Crozier, R. H. (2010). Integrační taxonomie: multisource přístup k zkoumání biologické rozmanitosti. Annu. Reverend Entomol. 55, 421–438. doi: 10.1146/annurev-takže-112408-085432
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Seehausen, O, Takimoto, G., Roy, D., a Jokela, J. (2008). Obrácení speciace a dynamika biodiverzity s hybridizací v měnících se prostředích. Mol Ecol. 17, 30–44. doi: 10.1111 / j. 1365-294X. 2007. 03529.x
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Pastýř, K. a., Thiele, K. R., Sampson, J., Coates, D. J., a Byrne, M. (2015). Vzácný, nový druh Atriplex (Chenopodiaceae) zahrnující dvě geneticky odlišné, ale morfologicky kryptické populace v suché západní Austrálii: důsledky pro taxonomii a ochranu. Aust. Syst. Bota. 28, 234–245. doi: 10.1071 / SB15029
CrossRef Full Text / Google Scholar
Singhal, s., and Bi, K. (2017). Historie čistí nepořádek: dopad času na divergenci a introgresi v tropické zóně šití. Evoluce 71, 1888-1899. doi: 10.1111 / evo.13278
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Singhal, S., Hoskin, C., Couper, P., Potter, S., a Moritz, C. (2018). Rámec pro řešení kryptických druhů: případová studie z ještěrek australských vlhkých tropů. Syst. Biol. . doi: 10.1093 / sysbio / syy
PubMed Abstract / CrossRef Full Text / Google Scholar
Singhal, s., and Moritz, C. (2013). Reprodukční izolace mezi fylogeografickými liniemi se stupňuje s divergencí. Proc. Roy. Soc B 280: 20132246. doi: 10.1098 / rspb.2013.2246026
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Steane, D. A., Potts, B. M., McLean, E., Collins, L., Prober se, S. M., Stock, W. D., et al. (2015). Skenování v celém genomu odhaluje kryptickou strukturu populace v eukalyptu přizpůsobeném suchu. Strom Genet. Genomes 11: 33. doi: 10.1007/s11295-015-0864-z
CrossRef Plný Text | Google Scholar
Udeřil, T. H., Feder, J. L., Bendiksby, M., Birkeland, S., Cerca, J., Gusarov, V. I., et al. (2017). Hledání evolučních procesů skrytých v kryptických druzích. Trendy Ecol. Evol. 33, 153–163. doi: 10.1016 / j. strom.2017.11.007
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Sukumaran, J., a Knowles, L. L. (2017). Multidruhové koalescence vymezuje strukturu, nikoli druh. Proc. Adresa. Aca. Věda. U. S. A. 114, 1607-1612. doi: 10.1073 / pnas.1607921114
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Swarts, N. D., Clements, M. a., Bower, C. C., a. Miller, J. T. (2014). Definování ochranářských jednotek v komplexu morfologicky podobných, sexuálně klamných, vysoce ohrožených orchidejí. Biol. Nevýhoda. 174, 55–64. doi: 10.1016 / j. biocon.2014.03.017
CrossRef Plný Text | Google Scholar
Tautz, D., Arctander, P., Minelli, A., Thomas, R. H., a Vogler, a. P. (2003). Prosba o taxonomii DNA. Trendy Ecol. Evol. 18, 70–74. doi: 10.1016/S0169-5347(02)00041-1
CrossRef Full Text | Google Scholar
Taylor, HR, Dussex, N., and van Heezik, y. (2017). Překlenutí mezery v genetice ochrany identifikací překážek implementace pro odborníky na ochranu přírody. Globus. Ecol. Konzervativec. 10, 231–242. doi: 10.1016 / j. gecco.2017.04.001
CrossRef Plný Text | Google Scholar
Thomson, S. a., Pyle, R. L., Ahyong, S. T., Alonso-Zarazaga, M., Ammirati, J., Araya, J. F., et al. (2018). Taxonomie založená na vědě je nezbytná pro globální ochranu. PLoS Biol. 16: e2005075. doi: 10.1371 / deník.pbio.2005075
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Vieites, D. R., Wollenberg, K. C., Andreonec, F., Köhler, J., Glaw F., Vences M. (2009). Obrovské podcenění biodiverzity Madagaskaru, o čemž svědčí integrativní soupis obojživelníků. Proc. Natle. Acad. Věda. USA 106, 8267-8272. doi: 10.1073 / pnas.0810821106
CrossRef Plný Text
Vogler, a. P., and de Salle, R. (1994). Diagnostika jednotek řízení ochrany. Nevýhoda. Biol. 8, 354–363. doi: 10.1046 / j. 1523-1739. 1994. 08020354.x
CrossRef Full Text / Google Scholar
Waples, R. S. (1991). Losos obecný, Oncorhynchus spp., a definice „druhů“ podle zákona o ohrožených druzích. Březen. Ryby. Rev. 53, 11-22.
Google Scholar
Waples, R. S. (1995). „Evolučně významné jednotky a zachování biologické rozmanitosti podle Zákona o Ohrožených Druhů,“ v Evoluci a Vodní Ekosystém: Definování Unikátních Jednotek v Populaci Ochranu, eds J. L. Nielsen (Bethesda, MD:Americké Rybářské Společnosti), 8-27.
Google Scholar
Waples, R. S., Gustafson, R. G., Weitkamp, L. a., Myers, J. M., Jjonson, O. W., Busby, P. J., et al. (2001). Charakterizující rozmanitost lososa z severozápadního Pacifiku. J. Fish Biol. 59, 1–41. doi: 10.1111 / j. 1095-8649.2001.tb01376.x
CrossRef Plný Text | Google Scholar
Waples, R. S., Nammack, M., Cochrane, J. F., a Hutchings, J. a. (2013). Příběh dvou aktů: praktiky seznamování ohrožených druhů v Kanadě a Spojených státech. BioScience 63, 723-734. doi: 10.1093 / bioscience / 63.9.723
CrossRef Full Text / Google Scholar
Waples, R. S., Punt, a. E., and Cope, J. M. (2008). Integrace genetických dat do řízení mořských zdrojů: jak to můžeme udělat lépe? Ryba 9, 423-449. doi: 10.1111 / j. 1467-2979. 2008. 00303.x
CrossRef Plný Text | Google Scholar
Wege, J. a., Thiele, K. R., Shepherd, K. a., Řezník, R., Macfarlane, T. D., a Coates, D. J. (2015). Strategická taxonomie v biodiverzní krajině: nový přístup k maximalizaci výsledků ochrany vzácné a špatně známé flóry. Biodivers. Konzervativec. 24, 17–32. doi: 10.1007/s10531-014-0785-4
CrossRef Plný Text | Google Scholar
Whitlock, R. (2014). Vztahy mezi adaptivní a neutrální genetickou rozmanitostí a ekologickou strukturou a fungováním: metaanalýza. J. Ecol. 102, 857–872. doi: 10.1111 / 1365-2745. 12240
PubMed Abstract / CrossRef Full Text / Google Scholar
Wiley, e.o. (1978). Koncept evolučních druhů se přehodnotil. Syst. Biol. 27, 17–26. doi: 10.2307 / 2412809
CrossRef Full Text / Google Scholar
Wilson, E.O. (2017). Výzkum biologické rozmanitosti vyžaduje více bot na zemi. Adresa. Ecol. Evol. 1, 1590–1591. doi: 10.1038/s41559-017-0360-y
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Yang, Z. H., a Rannala, B. (2014). Neřízené vymezení druhů pomocí sekvenčních dat DNA z více lokusů. Molo. Biol. Evol. 31, 3125–3135. doi: 10.1093/molbev/msu279
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Yeates, D. K., Seago, A., Nelsonová, L., Cameron, S. L., Josef, L., a Trueman, W. H. (2011). Integrační taxonomie, nebo iterativní taxonomie? Sys. Ent. 36, 209–217. doi: 10.1111/j.1365-3113.2010.00558.x
CrossRef Full Text | Google Scholar